原文链接:https://mp.weixin.qq.com/s/CSDIbiHxJCdvA6aYcXuj4Q
作者瞻云

(本篇8000字,综合大量文献研究,深度分析)
意识的本质是什么?它如何从物质的大脑里产生的?

人类尚没有确切的答案。
但或许,我们已经摸到意识世界的门槛。
本篇文章将整理主要的意识相关研究,试图得到一个比较清晰的脉络。
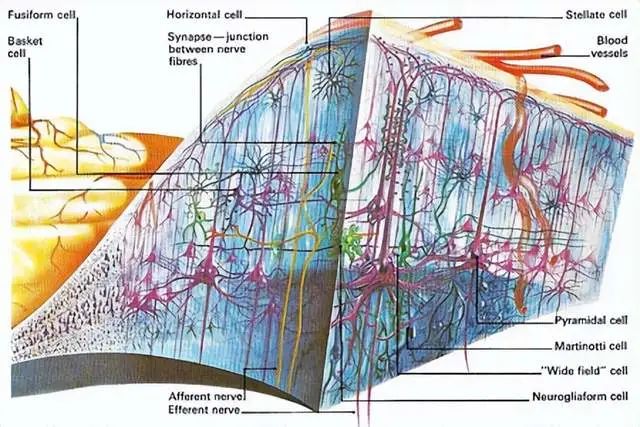
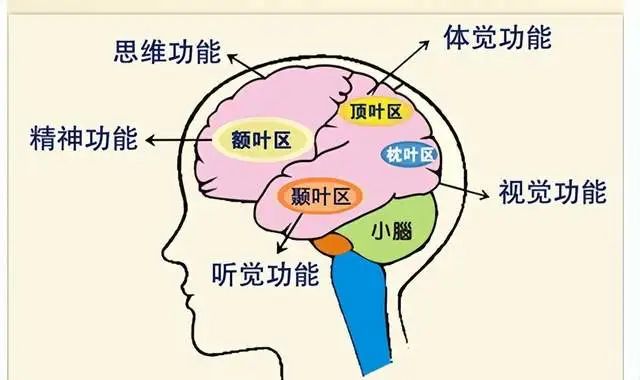
大脑皮层是人类最高级神经中枢,有着复杂的神经网络。
它一般被认为是人类意识的神经基础。
虽然,意识是什么至今没有定论,但通常情况下我们把意识到某种感觉/某个物体的存在,或者意识到我们行为/情感,判定是一种意识活动。
除了嗅觉(可直接透射边缘系统)外的绝大多数感觉,都会先投射到丘脑系统,经过初步整合,再通过丘脑皮层神经束投射到大脑皮层。

我们知道,大脑皮层功能是分区的:
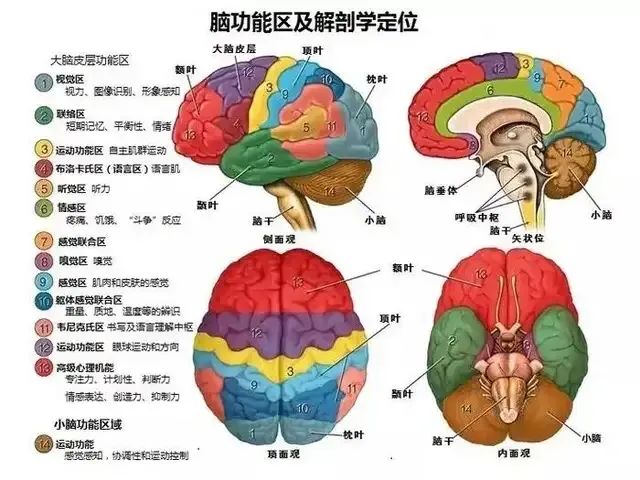
但当我们意识到一个物体时,它包含的信息却是复杂的。
例如,我意识到自己的手机放在桌上,它同时包含了手机的位置、大小、颜色,如果来了短信还会包含声音、画面等等。
也就是说,我们进行意识活动时,信息是高度整合的。
在我们正常认知活动的过程中,大脑暂时加工和储存的记忆被成为工作记忆。工作记忆的信息,随着时间的推移而被整合,诺贝尔生理学奖得主杰拉尔德·埃德尔曼认为,这个过程,可以在我们大脑中产生一个稳定的世界表象[1]。
大脑功能的贝叶斯方法,则对大脑多种感觉信息整合进行了成功预测[2][3][4]。
不同的大脑回路(例如,感觉系统、动作系统、语言系统等等)如何结合起来进行感知、决策和行动,产生明显统一性相协调的意识,被称为“绑定问题(Binding problem)[5]。
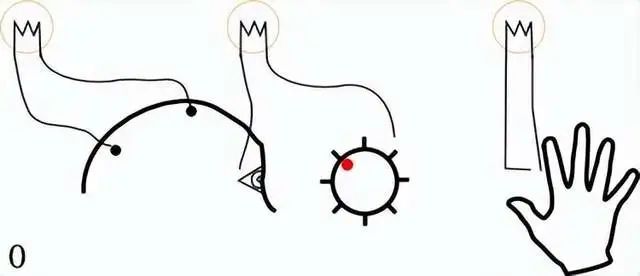
例如,以下四种图形,我们的大脑能够轻松地判断它们的区别:
一个红色的圆圈
一个蓝色的正方形
一个蓝色的圆圈
一个红色的正方形
大脑意识到这四种物体的区别,便是一种视觉特征绑定。
不难发现,绑定问题是意识问题的关键,如果我们能解决绑定问题,至少就已经敲开了意识世界的大门。
那么,我们的大脑又是如何统一我们所感知的世界呢?
我们知道,大脑皮层通过神经网络联络在一起:
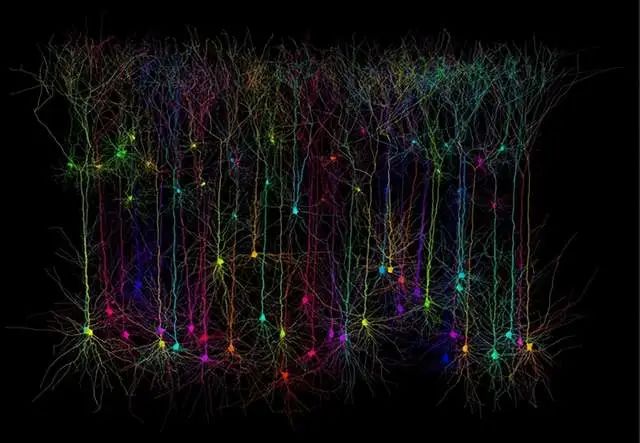
大脑皮层具有多层结构,仅仅2~4mm的新皮层,总共分成6层:
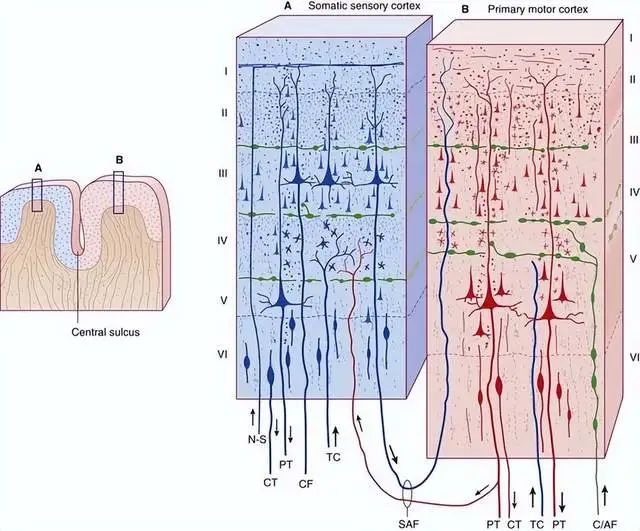
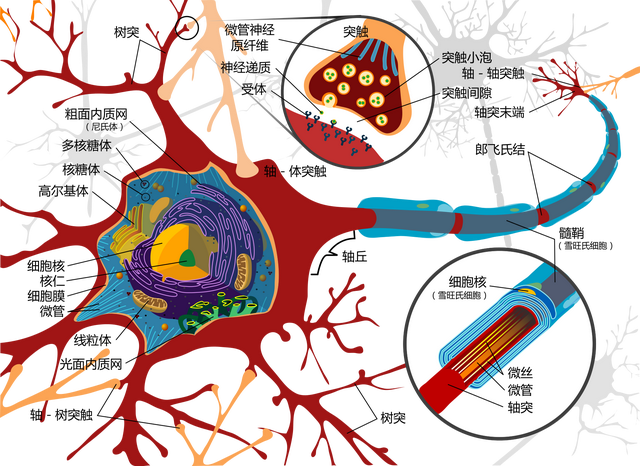
这6层结构中,主要存在锥体细胞、梭形细胞、颗粒细胞等三种神经元。
其中锥体细胞是大脑皮层所特有的,且是主要的投射(把信号发射到其它结构中去)神经元。
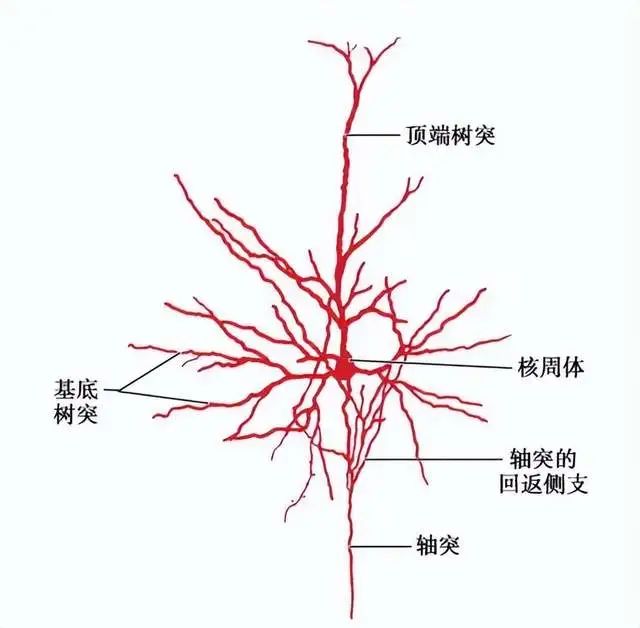
椎体神经元,具有丰富的树突,并通过轴突投射神经信号到其它脑区
虽然锥体细胞在不同的层级有不同的分布,但都有着丰富的树突结构,联络着整个大脑皮层。
尤其是第五层的大型锥体细胞,发出的神经纤维集合成束,往往能达到基底核、丘脑、延髓、脑干网状结构、脊髓等部位。
在大脑神经信号传递的过程中,神经元群体周期性的兴奋和抑制,形成特定节律的同步放电,从而产生神经振荡。
而神经振荡产生的电波便是脑电波。
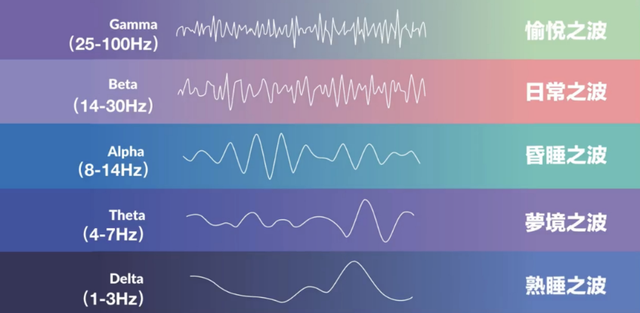
其中,25Hz以上的γ波,被证明与人的工作记忆、认知、注意力、感知、情绪相关,
而γ波,常见于丘脑皮层环路(丘脑向不同脑区投射信号,不同脑区也会向下投射信号)。
整个大脑皮层中有许多丘脑皮层环路,在有意识的感知过程中,皮层许多广泛分布的区域具有相互的同步通信模式。
在丘脑皮层环路中,不同大脑区域的γ波神经放电频率明显同步[6],警觉和专注度越高的动物,γ波也越突出[7]。当对单词进行感知时,大脑中的γ波也出现了跨皮质区域的同步振荡。
这些研究,不仅体现了意识活动的耦合性,也表明了意识活动与γ波的紧密关系[8]。
有研究者认为,这种支持远程大脑区域之间大规模同步事件的能力,可以提供连贯的感知[9][10]。
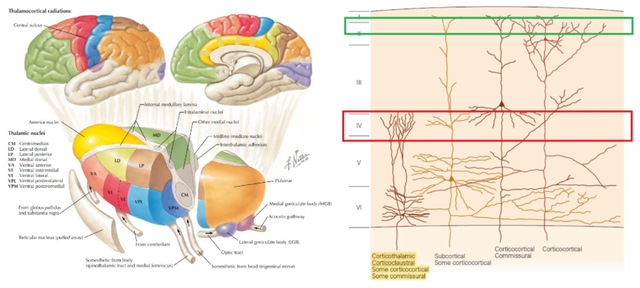
丘脑中央核团服务于唤醒和注意力,当丘脑受到一点点损伤,病人便会导致意识障碍[11],或陷入深度昏迷状态。
丘脑皮层神经环路对信息的整合处理,极可能是意识诞生的基础。
不仅哺乳动物都有这样的意识基础,就连缺乏新皮层的鸟类的意识基础,也在大脑皮层同源物中所识别[12]。
所以,丘脑皮层环路自然被认为有望解决绑定问题。
但要真正解决,依旧还需要弄清意识的真相。

有没有什么具体的实验,可以追踪意识活动的轨迹呢?
当然是有的。
最典型的就是双眼竞争。
双眼竞争:两个眼睛分别看不同物体时,视野会融合或互相替换[13]。
体验双眼竞争最简单的方式,是使用卷起的两个纸筒,分别用左右眼看眼前的物体(足够近,左右视野重叠),你会发现你视野中左右眼不同的画面也重叠在了一起。
看区别明显的画面,往往其中一个眼睛会更占优势。而看画面相近的,图像更容易发生融合。
例如,你左眼看键盘上的字母N,右眼看字母M,两个字母便会重叠起来。
如果左右眼看到的是没有冲突的两个画面,二者甚至可以比较完美的融合在一起。
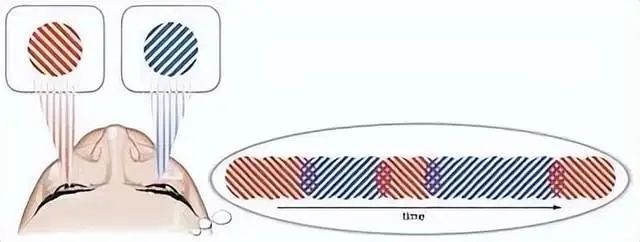
当双眼视觉交替竞争时,我们的意识感知也是交替发生的。
通过对猕猴进行研究发现,当出现双眼竞争时,大脑初级视觉皮层的活动基本没有什么变化,仅仅只有极少数神经元做出了相应的改变。相反,涉及到识别活动的高级中枢下颞叶皮层上,几乎所有的神经元都对优势视觉做出了反应[14]。
此类研究表明,大脑主要感觉区域的活动不足以产生意识[15]。而且在一项实验中,即使受试者大脑初级视觉皮层对刺激有明显的电反应,他们也缺乏意识感知。
近年来关于人类大脑神经振荡的相关研究也发明,丘脑自下而上的震荡活动,往往不容易产生意识,而高级皮层自上而下的活动,往往更容易产生意识[16]。
甚至当感觉信号与大脑内部的感觉意识不一致时,自下而上的反馈可以否决初级感觉皮层的活动,从而使其对感知不可见[17]。
虽然初级感觉皮层不容易单独产生感觉意识活动,但却会直接影响感觉意识的结果。
例如,当我们主观感受到更亮的光时,大脑内脑电频率的变化仅仅发生在初级视觉皮层内[18]。
也就是说感知本身不参与意识活动,但它却能决定意识活动中的感觉质量。
这样我们便能梳理出感觉意识的大致途径:
我们身上的各类感觉神经末梢,在一瞬间就能产生无比庞杂的感觉信息,这些感觉信息会通过丘脑进行初级的筛选和整合,这样会过滤掉大量无用的信息。
丘脑整合后,再通过丘脑皮质环路投射到初级感觉皮层。初级感觉皮层整理信息之后,通过皮层联络投射到更高级中枢。
更高级中枢可以选择性地提取这些信息,并让我们对特定的信息产生意识感知。
例如,当我在专注打字时,我的注意力在屏幕上,我不会有意识地去感知我接触地面的脚掌,在这一刻我也不会意识到任何的脚底感觉。
但我提到这一句话的时候,我却主观地去感知了脚底,并意识到我脚踩在地上,并可以一一去感知我脚底的各个部位。
来自脑干、丘脑、初级感觉皮层、边缘系统的各类信息,可能仅仅只是意识加工的原材料,初级中枢到高级中枢之间的联系,是意识产生的必然途径。
通过电刺激屏状体[19]、中央外侧丘脑[20]、脑干网状结构[21],都有发现意识关停现象,或许这些脑结构恰好是感觉、知觉到意识形成的关键通道。
大多失去意识的植物人,都有脑干网状结构受损,而屏状体和中央外侧丘脑都可以通过电刺激反复开关意识,屏状体功能发现较早,中央外侧丘脑是近年来的发现。
意识开关的存在,其实也是侧面证明了意识活动是多脑区的整合。
综合以上所有的研究来看,意识活动也更像信息处理之后的一种整合的感知。
然而,这种感知却是延迟的。
早在20世纪80年代,心理学家本杰明•李贝特便做了一个著名的实验[22][23]。
他们让5个左撇子的大学生坐在躺椅上,并告诉他们用1~2秒的时间放松头部、颈部,以及前臂肌肉。但在决定做这一件事情之前,他们需要突然快速动一根手指或手腕。当他们活动手指的时候,不要有任何的预先计划或刻意关注,随机重复40次。
在这些大学生进行这些动作时,研究人员测量了三个变量:
1、贴在前臂的电极,记录手指动作开始的时间。
2、贴在头皮上的电极,测试动作开始时大脑的预备电位。
3、感受到行为冲动(想动手指)时,喊出屏幕中钟表的“时间”,从而测出决定时刻。
经过多达几百次实验,李贝特最终都发现,决定时刻出现在大脑预备电位之后。
平均时间间隔为350ms。
也就是说,当我们决定做某一件事情之前的1/3秒,大脑就已经发起动作了。
因此,有研究者悲观地认为人类并没有自由意识。
不过在进一步的实验中,李贝特让被试大学生,在做决定之后否决行为。
虽然大脑出现了预备动作电位,但最终阻止了动作,没有检测到手上的电位。近年来的也有研究进一步表明,当大脑动作电位出现后一定时间内,可以进行否决,但距离动作时间足够近时,否决的成功率便会大大降低[24]。
这说明,无论人类有没有自由意识,但在一定时间内都有否决的自由。
40年来,有大量的研究支持李贝特实验中意识决定延于与大脑动作的结果。延后时间短则数百毫秒,最长甚至可达10余秒[25][26][27]。
一项实验中,经颅磁刺激改变受试者的左右手使用习惯后,受试者依旧认为自己的选择经过了自由意识的决定[28]。在某些实验设计下,当出现无意识判断或冲动行为,受试者也会认为是自己的决策行为[29]。
直接对高级皮层进行刺激,受试者则可能出现错误的意识判断。例如,他们可能认为自己做出了某种动作,但实际并非发生[30]。
这似乎更加肯定了人类没有自由意识。
然而,最近10多年一些研究者却有了进一步的发现,有研究者否定了人类没有自由意志的看法。

2009年,有人把李贝特的经典实验,修改为播放一段音频,然后让志愿者决定是否敲击一个键。研究发现,不管志愿者是否真的选择了敲击,两种情况下都有相同的大脑预备电位。
这表明,大脑预备电位并不表明已经做出了决定。
当然志愿者即刻决定是用左手还是右手按键时,大脑的早期动作电位同样没有什么区别,这说明大脑早期产生的动作电位,可能是注意到信号或者对信息的预处理。
近年来,越来越多更精确的方法,证明意识决定不是瞬间出现的,而是逐步建立起来的。因此有研究者认为,决策结果的早期神经标记不是无意识的,而是简单地反映了有意识的目标评估阶段,这些阶段还不是最终的,在达成最终意识活动之前,这个决策可以终止或改变[31]。
总之,一个动作可能在我们的“意识”意识到它之前就已经开始了,并不意味着我们的意识不能批准、修改或者取消这个动作。

结合丘脑皮质环路,初级皮层到高级皮层投射,我们不难得到这样的推测:
大脑整合信息本身就是先到初级皮层,然后再到更高级的皮层。初级皮层随时随地都在获得信息,以供高级皮层使用。高级皮层在做决定之前,总是先要调控初级皮层的信息。在获得信息之后,有决定去做或者不去做的权利。
而当初级皮层先对我们的躯体进行控制,信息反馈到高级皮层后,也可能被当做成我们的主观决定。
这样,大脑依旧是具有一定程度自由意识的。
当然“自由意识”概念的定义本身也是充满争议的,在对“自由意识”具有更严格定义的人眼中,人依旧是没有自由意识的。
无论意识是否自由,李贝特实验都证明了意识是大脑进行信息整合时产生的一种感知。
那意识究竟只有高等动物才有,还是大多数生物的共有特性?
开篇我们探讨了人类意识与丘脑皮层环路的关系,在以人为中心的研究者眼里,大脑皮层(尤其是新皮层)、丘脑皮层环路是产生意识的必要条件,因此他们认为新皮层很少,甚至没有的动物并不具备意识基础。
然而非脊椎动物和低等脊椎动物,与高等脊椎动物大脑(或中枢神经系统)之间,并非绝对的继承关系。因为它们没有丘脑皮层环路,在无完全根据的情况下,判断其他形式的意识产生条件并不存在,无异于是一种人类中心主义的傲慢。
更何况,意识的产生也是一个逐渐的过程。
我们假设1.8亿~3.5亿年间起源的(类)哺乳动物,具有真正意义上的意识,那么从进化的逻辑来说,在3.5亿年前的脊椎动物也应该是具有意识基础的。

持以此类观点的研究者并不在少数。
甚至有人认为,细胞中微管水平的量子效应与意识有关。这样,意识的起源甚至可以早到数十年前[32],当然这种观点并不被多数人支持。
前寒武纪时代两侧对称动物起源时,进化出了比刺胞动物更为复杂的梯形神经系统,有了联想学习和记忆能力,例如蠕虫和海兔身上的研究已经充分证明。因此有人认为,它们已经产生了意识[33]。
但计算机也有学习和记忆功能,但距离产生意识却十分遥远。
因此,这种观点也只有少数人支持。

早期脊椎动物是否能产生意识,则得到了更多人的关注。
有研究者提出,脊椎动物的初级意识早在寒武纪时期,就已经起源[34]。
所有的脊椎动物都有相同的神经系统渊源[35]。
虽然这个时期的动物不可能存在新皮层,但它们的大脑却有除嗅觉之外“高等”脊椎动物端脑结构。例如,用于选择和维持行为动作的纹状体,用于情绪的杏仁核和其他边缘结构,以及用于形成记忆、空间导航的海马。
在只有原始中枢神经的文昌鱼中,存在大量简单无意识反射(例如光动反应、反射性撤退)。面对复杂的环境,无意识的反射是机械的。寒武纪时期,早期脊椎动物生活在食物链的底层,这就迫使部分活下来的种群,感官中枢变得越来越复杂,并逐渐出现了整合各类感官,以应对复杂环境的能力。
中脑中有一个叫做四叠体(顶盖)的结构。
这个结构在人类大脑中仅仅只是视觉的中继站。
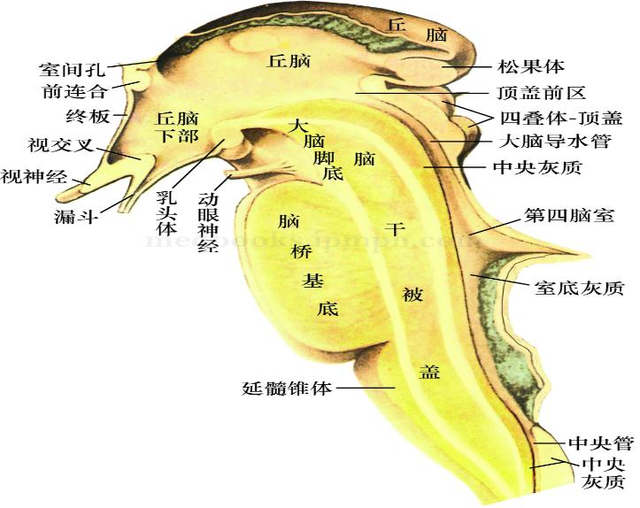
但它却是在非鸟和非哺乳脊椎动物脑中十分发达。
顶盖不仅是它们的主要感觉部位,它的层状组织,也能有效且广泛地整合来自不同感官的同构输入。
七鳃鳗是现存最为原始的脊椎动物,它们拥有真正的眼睛。
它们的大脑拥有视觉、嗅觉、体感和其他感觉的所有必要区域和神经分层路径。这些信息整合在一起产生注意力、知觉、神经同步和记忆——所有这些元素都被认为是意识形成所必需的。
研究者认为,丘脑-顶盖间的互相联系,可以多模态同构感觉表征,鳃鳗有至少感官意识。
当早期鱼类进化为合弓纲(类哺乳动物)和蜥形纲爬行动物 ,背侧大脑皮层逐渐增大,并成为感觉意识的主导中心。
对于鸟类和哺乳动物来说,感觉意识的中心从视顶盖转移到了大脑皮层,从而扩展和丰富意识体验。

纵观所有的研究,我们不难发现,意识存在的最核心基础是感官的整合。
只要感官系统足够的发达,并拥有整合能力的脑结构,那就有诞生的意识的条件。大脑皮层可能是一个充分条件,而非必要条件。意识能力的高低,则可能同时取决于感官和整合能力。
除了脊椎动物外,节肢动物和软体动物的意识研究也并不多,但从信息整合的角度,我们或许有更多的启迪。
最后再回到人类意识的角度,谈一谈意识最可能的理论基础。

既然大脑是整合的,那么大脑最基本的意识单元是什么?
我们知道,单个感觉神经元的活动是十分嘈杂(具有很高的神经元噪声),无法在大脑内重建感觉场景。
为了解释大脑内的神经回路,早在1949 年,著名神经科学家唐纳德·赫布提出的赫布理论(突触可塑性的基本原理)中,便有了神经元集群的概念。
相关实验也表明,当猴子进行伸手和抓握运动时,神经元集群同时编码手臂位置、速度和手的抓握力,注意和记忆的位置都可以被解码的。[36]
到了20世纪末,有神经科学家通过神经元群体解释了运动皮层的编码方向。
在神经元集群的概念下,其实大脑神经通路和稻草编制的长绳很相似,你追溯单根稻草的轨迹是很难得到整根绳子信息的,所有稻草信息的集合才是整根绳子。单根稻草的不确定信息在集合成整根绳子的时候,被平均掉了,最终得到的是整根绳子的信息。
神经元集群的基本单位是皮质柱(Cortical Column)[37]。
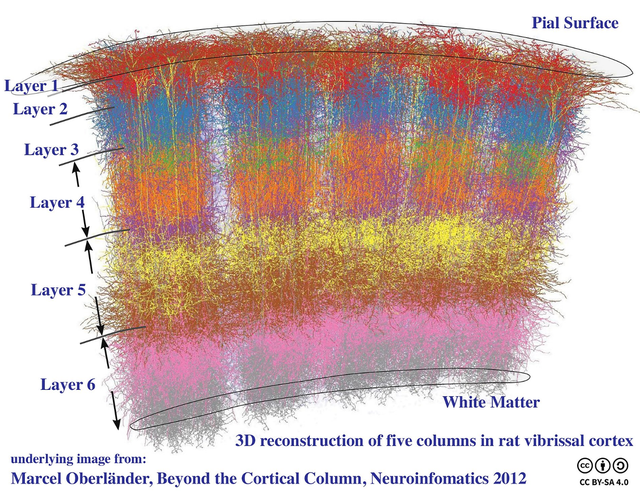
通过在皮质表面垂直插入探针连续穿透,发现的几乎相同的感受野(receptive field,一个神经元所反应的刺激区域),因为把整个区域的神经元集群称为皮质柱。
皮质柱内的神经元编码具有相似特征,这支持了大脑皮层的模块化。
虽然皮质柱假说依旧是当前解释皮层信息处理的最广泛假说之一[38],然而遗憾的是,模块化的功能结构和遗传机制,依旧没有相关研究结果所支持[39]。
从皮质柱到意识产生,此中有没有什么基本的意识框架呢?
虽然没有实锤的证据,但当前主流的有四大意识理论[40],我这里再简单谈谈。

1、高阶理论(HOTs)
心理状态→元心理状态(高阶表象)
该理论认为,普通心理状态转变成高阶的元心理,从而诞生了意识。
一般来说,前额叶和人类的高级认知功能密切相关[41]。
前额叶皮层分为背外侧、腹外侧、内侧和眶额皮层,各个区域有不同的功能,但并不是这里分析的重点[42]。
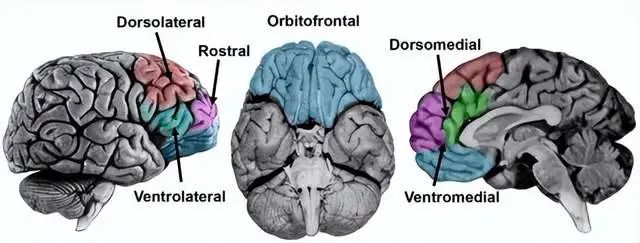
前额叶最晚进化出现,也是人类发育最晚成熟的脑结构,是人类最高级中枢,执行着抽象认知、注意力调控、行为决策、思维推理、工作记忆等与意识高度相关的活动。
高阶理论其实是建立在前额叶功能的基础上的,但个人认为这还是过于形而上。
作为高阶的元心理是凌驾于普通心理之上的,而前额叶和其它高级皮层终究是并列的,且并不仅仅是前额叶参与意识的形成,其它一些关键脑区同样参与。
但如果我们定义前额叶诞生的意识就是元心理,这个理论便陷入循环论证。
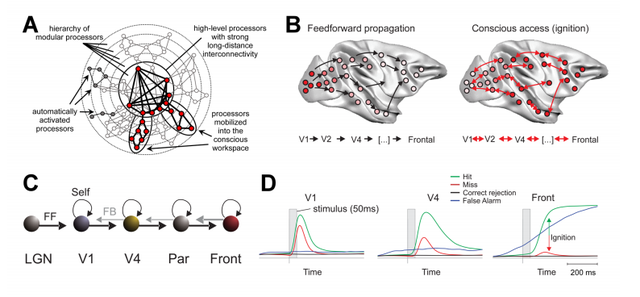
全局工作空间理论(GWT)
这个理论,是把大脑看做是执行不同功能的专用模块,并通过长距离连接,信息在不同模块之间传播和共享,每一时刻的及时全局共享诞生意识。
这个理论很明显是以大脑分区以及联络关系作为理论基础的,它同样有脑科学的基本研究支撑。但这个理论关于全局的概念,依旧是抽象而不明晰的。
如果我们定义能诞生意识的共享就是全局的,这个理论同样会陷入循环论证。
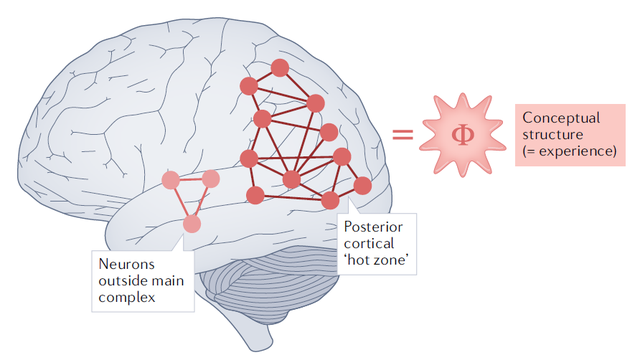
信息整合理论(IIT)
这个理论明显是以大脑的信息整合机制作为基础的,它意图以现象特征为基础,构建一套几何式公理,显示出了庞大的野心 。
该理论的基础逻辑,是探讨大脑整合信息时的因果机制和状态。它提出,任何系统,只要产生一个非零的不可约的整合信息的最大值,便足以诞生仪式。
一直以来,我都认为信息整合理论是解释意识起源的最可能的方向之一。但个人认为,这个整合过程也可能是混沌的,它是否真的存在一个标准范式,是值得争议的。而且,即便真的存在标准范式,意识的诞生方式,也并不一定是该理论提出的形式。
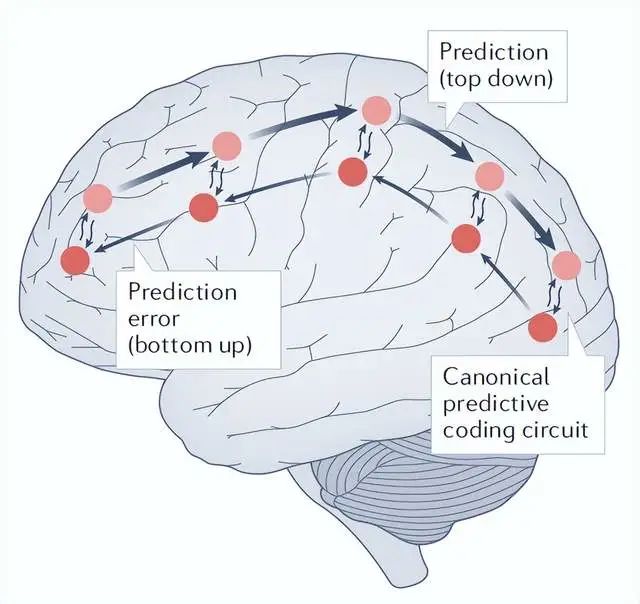
再入和预测处理理论
该理论强调了自上而下信号对形成和塑造有意识感知的重要性。
大脑(高级皮层)再处理(或再发生)感知皮层内的局部信息便足以产生意识。
这个理论,同样脱胎于现有的意识相关研究。
似乎能很好地解释一些意识研究自上而下的机制,以及关于意识的延迟现象。
这个理论的确能够解释局部意识,相当于大脑对感官产生原因的最佳猜测。
除了以上四个理论,还有一个理论的呼声很大,那就是“注意力图式”理论。
该理论的核心主张是,“注意力图式”是大脑计算注意力过程和当前状态的简化模型[43]。
根据该理论,诸如“我意识到XX”包括以下步骤:
刺激XX在大脑中被编码为一种表征,与其他刺激表征竞争大脑有限的处理资源。
如果刺激XX赢得了这种信号竞争,它就会被大脑深度处理,从而加入“注意力图式”。
“注意力图式”是一种内在模型,是选择性地获取XX的一些基本特征,例如颜色、形状、位置等,从而诞生的一种简化模型。
其实我们不难发现,当我们意识到一个物体时,当把这个物体拿走,我们也能在脑海中构想出它的基本特征。虽然我们无法确定“注意力图式”理论的正确性,但大脑处理工作记忆的方式,很明显是符合“注意力图式”。
“注意力图式”是最为机械化解释意识的一种理论,但再结合以上四种意识理论,我们或许更加接近意识的真相。
首先,意识是整合的,高级皮层再处理不同分区初级皮层中的信息时,产生的一种简化模型,可能是意识形成的基础。这个整合过程,或许是通过某种范式(例如数学关系)达成的。如果意识过程超出一般心理的范畴,它也而可能是更高阶的。
最后,关于意识的提取问题。
意识终究是某种信息,无论脑电波破译技术的突破,还是本身提到的诸多意识研究来说,意识当然是可以通过技术手段读取的。
但目前的能力,仅仅是读取某些特征信息,对于整个意识依旧是困难的。
要真正的做到完全的读取意识信息,即便不需要意识理论的完全建立,也需要建立一个十分近似模型,来进行读取。
退一万步说,哪怕人类永远得不到意识的真相,但只要我们的模型无限接近意识特征,我们未来读取的意识,就能无限接近我们的真实意识。
当然,从意识诞生的可能机制来看,意识上传可能是一种永远不可能达成的幻想。

◆写在最后◆
本篇文章花了好几天的时间来撰写,意识的主要研究应该都有所呈现。
可以看出,不同的研究者对意识的看法不尽相同,有的甚至完全相反。
其实,关于意识的定义,在各类心理学、神经科学专著,以及词典中的定义都不尽相同。
有的定义可能更偏初级感知,有的可能更偏高级知觉,注意力,甚至决策,也有的都囊括在了其中。
为了避免争议,本篇中的意识,主要是指“意识到XX”的一种感知或注意。
本篇除了相关研究和研究者的观点以外,也在一些地方明确提出我自己的看法。
关于各种研究、推论,以及假设,需要辩证来看待。
◀喜欢本篇,点赞关注▶
《参考文献》
[1] Stafford, Calvin. The Remembered Present: A Biological Theory of Consciousness[J]. Jama the Journal of the American Medical Association, 1990, 264(13):1758.
[2] Battaglia P W , Jacobs R A , Aslin R N . Bayesian integration of visual and auditory signals for spatial localization[J]. Journal of the Optical Society of America A, 2003, 20(7):1391-1397.
[3] Knill, D, C. Learning Bayesian priors for depth perception[J]. Journal of Vision, 2010.
[4] Daniel G , Tong J . Prediction, Postdiction, and Perceptual Length Contraction: A Bayesian Low-Speed Prior Captures the Cutaneous Rabbit and Related Illusions[J]. Frontiers in Psychology, 2013, 4:221-.
[5] Feldman J . The neural binding problem(s)[J]. Cognitive Neurodynamics, 2013, 7(1):1-11.
[6] Buzsaki G . Rhythms of the Brain[M]. 2009.
[7] Mccormick D A , Mcginley M J , Salkoff D B . Brain state dependent activity in the cortex and thalamus[J]. Current Opinion in Neurobiology, 2015, 31:133-140.
[8] Melloni, Lucia et al. “Synchronization of neural activity across cortical areas correlates with conscious perception.” The Journal of neuroscience : the official journal of the Society for Neuroscience vol. 27,11 (2007): 2858-65. doi:10.1523/JNEUROSCI.4623-06.2007
[9] Schiff N D . Central Thalamic Contributions to Arousal Regulation and Neurological Disorders of Consciousness[J]. Annals of the New York Academy of Sciences, 2008, 1129(1):105-118.
[10] Edelman G M , Gally J A , Baars B J . Biology of Consciousness[J]. Frontiers in Psychology, 2011, 2.
[11] Boly M , Seth A K . Modes and models in disorders of consciousness science.[J]. Archives Italiennes de Biologie, 2012, 150(2/3):172-184.
[12] Rose J , Gunturkun O N , Kirsch J A . Evolution of Association Pallial Areas: In Birds. Springer Berlin Heidelberg.
[13] Blake, R., Logothetis, N. Visual competition. Nat Rev Neurosci 3, 13–21 (2002).
[14] Logothetis N. (1998) Single units and conscious vision. Philos. Trans. R. Soc. Lond. B, 353: 1801–18.
[15] Owen A . The Quest for Consciousness[J]. 2011.
[16] Crick, Francis, Koch, et al. A framework for consciousness.[J]. Nature Neuroscience, 2003.
[17] Li Z . A new framework for understanding vision from the perspective of the primary visual cortex. 2019.
[18] Bie De Rlack J , Castelobranco M , Neuenschwan De R S , et al. Brightness induction: rate enhancement and neuronal synchronization as complementary codes.[J]. Neuron, 2006, 52(6):1073-1083.
[19] Crick F C , Koch C . What is the function of the claustrum?[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2005.
[20] Redinbaugh M J , Phillips J M , Kambi N A , et al. Thalamus Modulates Consciousness via Layer-Specific Control of Cortex[J]. Neuron, 2020, 106(1).
[21] Waldemar Iwańczuk, Guniczak P . Neurophysiological foundations of sleep, arousal, awareness and consciousness phenomena. Part 1.[J]. Anaesthesiology Intensive Therapy, 2015, 47(2):168-174.
[22] Libet B , Gleason C A , Wright E W , et al. TIME OF CONSCIOUS INTENTION TO ACT IN RELATION TO ONSET OF CEREBRAL ACTIVITY (READINESS-POTENTIAL)[J]. Brain, 1983, 106(3):623-642.
[23] Libet B . Unconscious cerebral initiative and the role of conscious will in voluntary action[J]. Behavioral & Brain ences, 1985, 8(4):529-539.
[24] Schultze-Kraft M , D Birman, Rusconi M , et al. The point of no return in vetoing self-initiated movements[J]. Proc Natl Acad Sci U S A, 2016, 113(4):1080-1085.
[25] Matsuhashi M , Hallett M . The timing of the conscious intention to move[J]. European Journal of Neuroence, 2010, 28(11):2344-2351.
[26] Soon, C., Brass, M., Heinze, HJ. et al. Unconscious determinants of free decisions in the human brain. Nat Neurosci 11, 543–545 (2008).
[27] Soon C S , He A H , Bode S , et al. Predicting free choices for abstract intentions[J]. Proceedings of the National Academy of Sciences, 2013, 110(15).
[28] Brasil-Neto J P , Pascual-Leone A , Valls-Sole J , et al. Focal transcranial magnetic stimulation and response bias in a forced-choice task.[J]. J Neurol Neurosurg Psychiatry, 1992, 55(10):964-966.
[29] Kühn, Brass M . Retrospective construction of the judgement of free choice[J]. Consciousness & Cognition, 2009, 18(1):12-21.
[30] Desmurget, M.; Reilly, K. T.; Richard, N.; Szathmari, A.; Mottolese, C.; Sirigu, A. (2009). "Movement Intention After Parietal Cortex Stimulation in Humans". Science. 324 (5928): 811–813.
[31] Guggisberg A G , Ana?S M . Timing and Awareness of Movement Decisions: Does Consciousness Really Come Too Late?[J]. Frontiers in Human Neuroscience, 2013, 7.
[32] Hameroff S . Did consciousness cause the cambrian evolutionary explosion?[J]. MIT Press, 1998.
[33] Ginsburg S , Jablonka E . The evolution of associative learning: A factor in the Cambrian explosion[J]. Journal of Theoretical Biology, 2010, 266(1):11-20.
[34] Feinberg, Todd E, and Jon Mallatt. “The evolutionary and genetic origins of consciousness in the Cambrian Period over 500 million years ago.” Frontiers in psychology vol. 4 667. 4 Oct. 2013.
[35] Baker C . Neural Crest and Cranial Ectodermal Placodes[J]. Springer US, 2005.
[36] Lebedev M A , Messinger A , Kralik J D , et al. Representation of Attended Versus Remembered Locations in Prefrontal Cortex[J]. PLoS Biology, 2004, 2(11):e365-.
[37] Buxhoeveden D P , Casanova M F . The minicolumn hypothesis in neuroscience[J]. Brain, 2002(5):935-951.
[38] Javier D F , Henry M , Rockland K S . The Neocortical Column[J]. Frontiers in Neuroanatomy, 2012, 6:162-173.
[39] orton J C , Adams D L . The cortical column: a structure without a function.[J]. Philosophical Transactions: Biological Sciences, 2005, 360(1456):837-862.
[40] Anil S and Tim B. Theories of consciousness. Nature Reviews Neuroscience, 2022.
[41] Cohen E H , F Tong. Neural Mechanisms of Object-Based Attention[J]. Cerebral Cortex, 2015(4):1080-92.
[42] Szczepanski SM, Knight RT: Insights into human behavior from lesions to the prefrontal cortex. Neuron 2014, 83:1002-1018.
[43] Graziano MS, Webb TW. The attention schema theory: a mechanistic account of subjective awareness. Front Psychol. 2015;6:500. Published 2015 Apr 23.
分享到朋友圈
深耕生命科学领域,探索宇宙万物之秘点击下方,关注瞻云
