原文链接:https://m.thepaper.cn/baijiahao_19691485?sdkver=93de1a00
原创 新宇 神经现实

文献:Luppi, Andrea I., et al. "A synergistic core for human brain evolution and cognition." Nature Neuroscience 25.6 (2022): 771-782.
DOI:10.1038/s41593-022-01070-0
作者:新宇 | 封面:Gerd Altmann / Pixabay
认知神经科学领域中,非常基本的一个理论假设就是把人类大脑看作一个分布式的信息加工系统。在大脑加工系统中,通过信息的传递来实现人的认知功能。以视觉信息为例:当我们用双眼观察世界的时候,两只眼接收的颜色信息在很大程度上都是冗余(redundant)的,但在深度的判断上则需要基于双眼感知到的第三个维度的差异信息来进行协同(synergistic)或互相补充。这两类信息都很重要,因为同时接受这两种信息意味着,大脑可以在利用冗余信息来维持鲁棒性(robustness)的同时,通过信息的整合(integration)来实现更高效的认知加工。所以,理解人脑认知加工过程中如何对这两种信息进行权衡(trade-off)将有助于洞悉人脑信息加工系统的架构。
因此,研究者试图解析大脑对于冗余信息和协同信息的加工。在传统的、基于功能连接(functional connectivity, FC)的脑网络框架中,很难区分和捕捉这两类信息。在本研究中,研究者利用了一种全新“信息分解”(information-resolved)计算框架来分解BOLD信号中的内在信息流,从而得到脑区在信息加工中起到的协同作用与冗余作用。这种方法被称为整合信息分解(Integrated Information Decomposition),它将大脑视为一个由脑区的当下状态以及脑区间交互来影响后续时序变化的动态系统。其中,冗余信息在当前状态中的每个脑区中都是均等可用的,而协同信息则必须从脑区间交互中(即通过信息的整合加工)额外产生。在此基础上,研究者试图解析其背后的组织模式与神经生物学基础。通过比较人类和非人灵长类(猕猴),研究者进一步阐明协同信息加工在人类复杂认知能力中的作用及其进化过程。

协同与冗余信息加工的脑组织模式及认知关联
更具体地,研究者采用了互信息(mutual information)来计算每个脑区的角色。香农的互信息(Shannon’s mutual information)定量描述了两个随机变量(X和Y)间的互相依赖程度。它刻画了源变量X可以在多大程度上为目标变量Y提供信息。部分信息分解(Partial information decomposition, PID)扩展了香农的理论,将X和Y都作为源变量,来衡量它们对于第三个变量Z的提供的信息量。这些信息可以被分解为不同的类型:只能由一个信息源提供,而另一个信息源不能提供(独特的信息);由两个信息源分别都可以提供(冗余信息);以及必须将两个信息源结合才能提供(协同信息)。整合信息分解在时间维度上进一步扩展了这一原理。与此同时,利用延时互信息(time-delayed mutual information,TDMI)可以用于评估整个系统过去到未来的信息流动总量。二者相结合,就可以将TDMI分解为对应于不同时间和不同变量间的三类变量。本研究中主要关注在时间维度上稳定出现的冗余信息和协同信息。
研究者从HCP中选取了100名健康成年被试的静息态功能MRI脑影像,获得了232个皮层上脑区与皮层下脑区的功能活动时间序列。通过整合信息分解,可以定量计算得到不同的脑区对在多大程度上参与冗余信息或协同信息加工,结果如图1所示(a和b)。通过将其与传统的FC进行比较(皮尔逊相关),可以发现FC与冗余加工的相似性要显著的高于协同加工,说明时间序列的同步性在很大程度上只能提供冗余信息而非协同信息。理论上,冗余加工和协同加工并不是一定呈现相反的模式,脑区间可以同时存在两种加工模式。
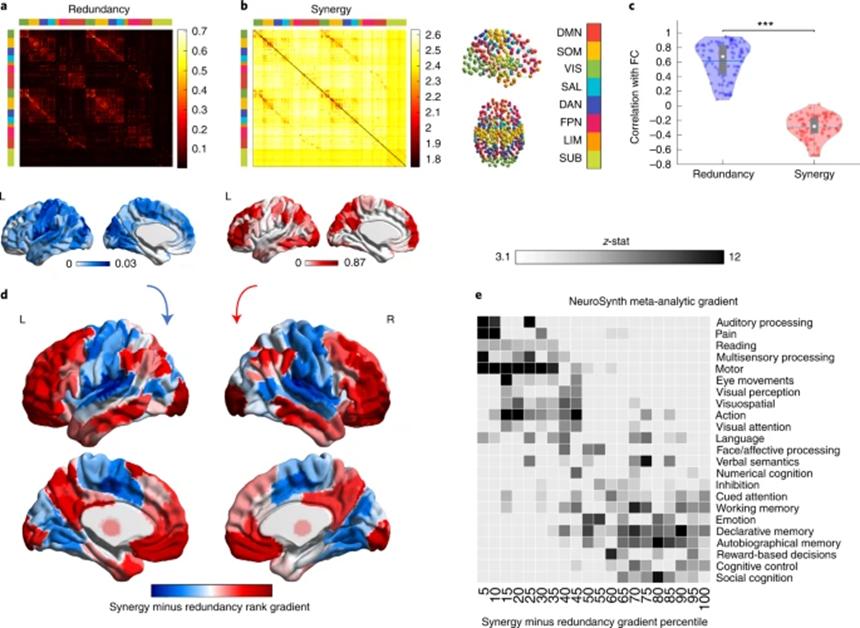
图1. 协同与冗余加工区域及其认知关联。
—
Luppi et al., Nat. Neurosci. (2022)
研究者进一步计算每个脑区与其他脑区间的相互协同高于相互冗余的程度(协同-冗余),来提供一个相对于冗余的协同程度参考,然后根据其进行了排序,得到了在更倾向冗余到更加倾向协同这一维度上存在梯度变化的脑区分布图(图1d)。根据Yeo的子网络划分,感觉运动区、凸显网络和大部分视觉区存在很高程度的冗余加工;对应于冯·艾克诺默(Von Economo)的细胞构筑分类,这些冗余信息加工区域主要集中在初级感觉、初级运动和岛叶皮层(如图2所示)。相反,在高级皮层脑区的协同程度要高于冗余,包括默认网络(DMN)、额顶执行控制网络(FPN)和边缘网络(眶额皮层和颞极等)。
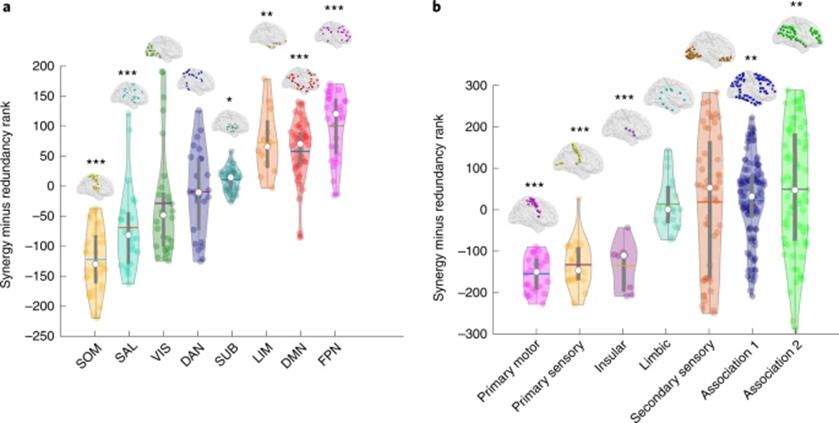
图2. 不同网络划分中的协同-冗余梯度模式。
—
Luppi et al., Nat. Neurosci. (2022)
冗余到协同的梯度模式与功能网络划分与细胞构筑层级分布的一致性,也提示了二者的交互可能在不同的认知领域存在差异。因此研究中利用NeuroSynth进行了基于术语的元分析,比较了24个认知术语(包括从低级感觉运动到高级的认知功能)的任务激活元分析结果与冗余-协同梯度空间模式的相似性。结果表明,高冗余的区域主要参与到听觉、视觉、多通道感觉加工和运动中;高协同的脑区则参与到社会认知、记忆和认知控制中,这些高级认知功能需要更复杂的信息整合(图1e)。
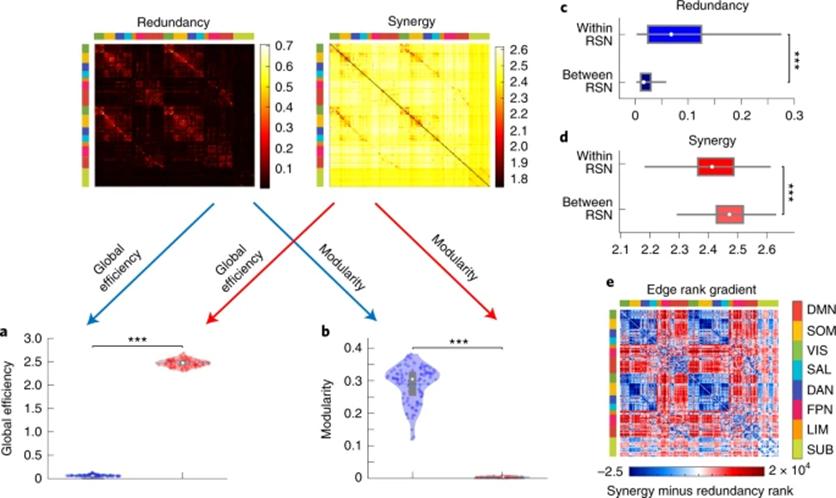
图3. 协同与冗余加工与脑功能网络拓扑属性的关联。
—
Luppi et al., Nat. Neurosci. (2022)
协同与冗余信息加工与功能网络拓扑及结构连接布线的关联
一种理论观点认为,认知架构被强制设计为能够同时满足两种不同且相反的需求:初级感觉运动需要将整个网络分为一些互相分离的模块,而高级认知功能的信息整合则需要一个高度连通的网络组织。本研究通过直接比较图论指标与冗余/协同的关系,来试图解答人脑如何解决上述特异性加工与全局整合间的矛盾。通过计算被试间的相关,研究者发现功能网络的全局效率与脑区间的协同的相似性要显著高于与冗余的相似性;相反地,网络的模块化指数则与脑区间的冗余信息的相似性显著高于与协同的相似性(图3)。因此这两种不同的信息交互可以用于反应两种存在明显区别的图论属性。进一步分析发现,冗余信息更多地存在于静息态功能子网络的内部而非网络间,同样地对于协同信息来说模式正好完全。说明在分离的子网络内的脑区间交互更依赖于冗余信息,而与此同时,协同信息则支持了不同子网络间的整合加工。
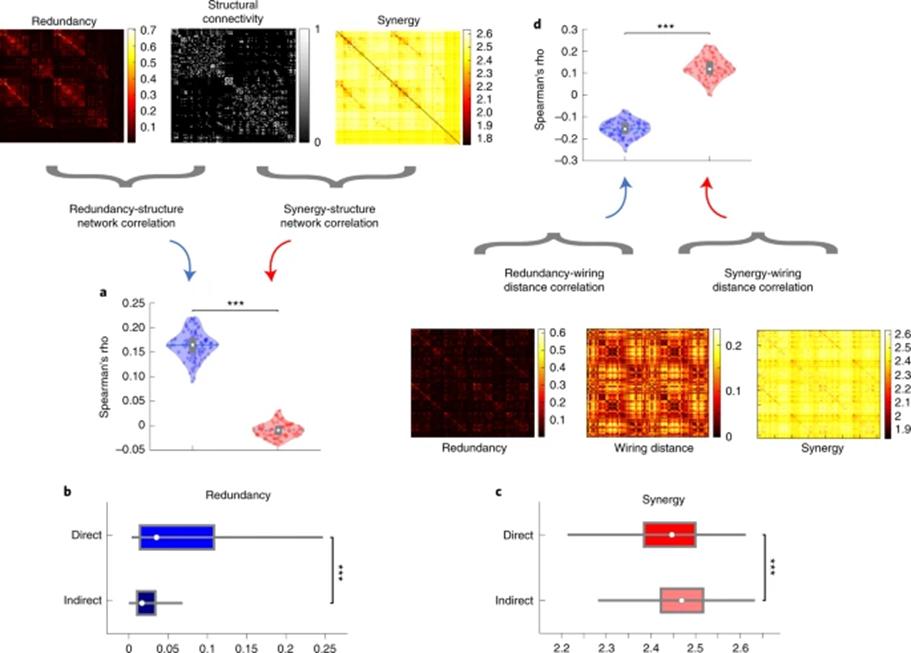
图4. 脑白质结构连接对于协同与冗余加工的支持。
—
Luppi et al., Nat. Neurosci. (2022)
为了明确信息交互是否存在一定的结构基础,研究者利用脑区间的结构连接来探索其与不同类型的信息交互之间的关系。因为对冗余信息的加工可以体现整个系统的鲁棒性,因此更应该存在直接的结构连接,这些结构连接才能实现脑区间最为可靠的沟通。结果表明,脑区之间的白质纤维数目与冗余信息交互在被试间存在较高的相似性,且显著地高于其与协同间的相关(如图4)。通过将脑区间的冗余性根据是否存在结构连接分为两类,进行组间比较的结果也验证了上述假设。对于协同信息,研究者推测,一方面如果脑区接收到了大脑中其他部分更丰富的输入,那它们会表现出更高的协同程度;另一方面,相似的信息输入常常会更多出现在冗余信息中。研究者为了验证这个假设,计算了脑区间的多尺度布线距离(multi-scale wiring distance),它结合了空间临近性(测地距离)和微结构相似性。结果表明,脑区间的多尺度布线距离与协同信息的相似性要显著的高于冗余信息,即连接模式差异越大的脑区对更加倾向于信息的协同交互。
简言之,在人类大脑中冗余信息的交互的划分出了一个模块化的结构-功能主干,确保了感觉运动的输入-输出通道的鲁棒性;而协同信息的交互则受益于结构连接模式的差异,实现了全脑不同子网间的高效连接,从而有助于高级认知加工。
协同信息加工的人脑遗传演化起源,及其在突触、代谢和分子水平的神经机制
为了实现独特的复杂认知能力,人脑需要信息高度的协同性。研究者认为协同信息加工为高级认知功能提供了可能性,因此汇集了3种方法来对这一假设进行验证。首先,研究者比较了人脑和非人灵长类(猕猴)的不同类型信息各自在总信息流中所占的比例。在冗余信息上,两者没有显著差异;但在协同信息加工上,人类的协同信息要显著的高于猕猴。将冗余-协同梯度变化呈现在猴脑上时(如图5c),可以观察到其整体模式与人类相似,体现出在演化中的相对稳定性。但在猕猴的前额叶皮层(PFC)中冗余信息的加工占主导地位,而该区域是在人脑中协同信息加工最强的区域。有趣的是,PFC也是在人类演化中存在实质性皮层扩张的区域。这一发现提示了,在人脑中较高的协同信息加工的区域分布可能与演化过程中的皮层扩张存在关联。因此研究者接着计算了冗余-协同梯度变化与人类相较于黑猩猩的皮层扩张脑图间的空间分布相似性。结果得到了显著的正相关,意味着演化中出现的额外皮层组织可能有助于协同信息加工的出现。最后,研究者利用艾伦脑科学研究院(Allen Institute for Brain Science)提供的脑遗传图谱,计算发现冗余-协同梯度模式与区域上特定的基因表达模式显著相关,包括与脑发育、脑功能(智力和突触传导)有关的基因,以及基因中与人脑相关的人类快速演化区域(HAR-Brain genes)。换言之,在人类中具有特殊功能的基因更倾向于表达在协同脑区。
总的来说,上述这些发现表明,人脑演化可能导致了信息协同交互的主导。
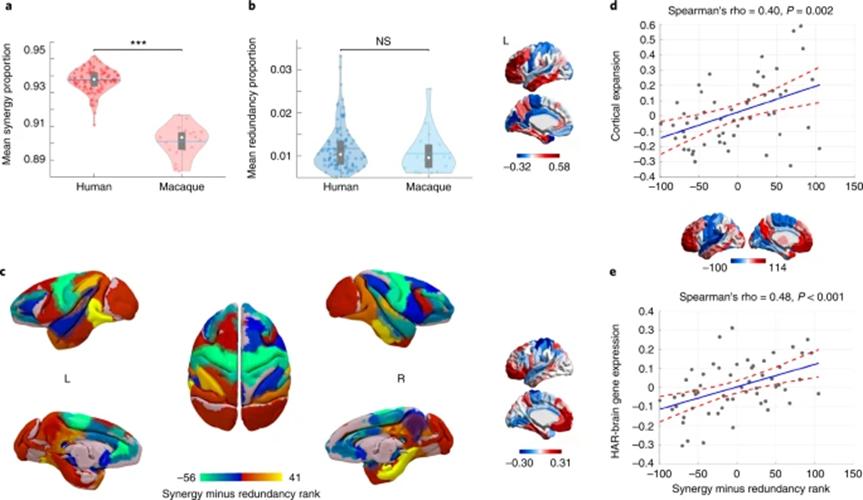
图5. 人脑演化中对协同信息加工的倾向。
—
Luppi et al., Nat. Neurosci. (2022)

图6. 人脑协同信息加工的突触相关神经机制。
—
Luppi et al., Nat. Neurosci. (2022)
人脑中如此高的协同加工是如何获得的?为了从神经生物学角度回答这一问题,研究者进一步对冗余-协同梯度与AIBS中提供的20674个基因的区域表达模式间的相似性进行了关联分析。利用偏最小二乘(partial least-squares,PLS)回归,得到了两个可以解释30%梯度变异的PLS成分(显著高于随机预期)。两个成分都显著地富集于基因的人类快速演化区域(HAR-Brain),与上面假设驱动得到的结果一致。通过基因本体(Gene Ontology,GO)分析得到成分的转录特征显著地富集于与学习记忆、突触、突触传导等相关的基因(图6a-e)。
突触是神经元之间信息交换的关键结构,因此它们组成了协同信息交互的最主要神经生物学基础。为了得到更直接的突触密度与协同区域主导性的关联,研究者利用正电子发射断层扫描(PET)在活体人脑中计算了区域的神经元密度。基于15个健康被试的独立数据,研究者取得了[11C]UCB-J PET结合电位的区域数值,可以将其分解为两个反应突触密度的全脑梯度分数主成分。结果表明,突触密度在全脑呈现的前后梯度变化的第二个主成分与冗余-协同梯度存在显著的相关(图6h)。
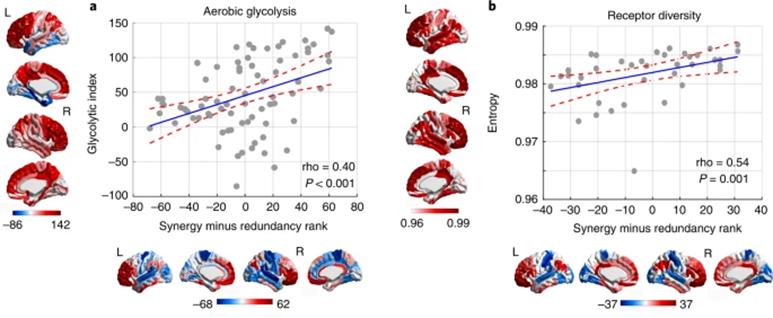
图7. 人脑协同信息加工的代谢与分子水平相关神经机制。
—
Luppi et al., Nat. Neurosci. (2022)
最后,研究者对协同信息加工的代谢基础和分子机制进行了探索。尽管上述的遗传和活体数据的结果已经表明了突触在协同信息加工中起到的作用,但突触本身并不是固定不变的,而是存在一定的空间和时间上的发育的。其中有氧糖酵解(aerobic glycolysis,AG)代谢过程可能支持了突触的生长与形成。研究者计算了反映有氧糖酵解能力的升糖指数(glycolytic index,GI),即PET测量的大脑中的氧和葡萄糖的代谢率。结果表明,升糖指数在全脑的空间分布模式与冗余-协同梯度存在显著的相关。
此外,尽管突触的密度可以反映整合多种信息的潜力,但神经元之间的实际交流需要多种不同的神经递质作用在多样的受体上。神经递质的分布在人类大脑的不同区域和相同区域的不同层(外颗粒层、颗粒层和内颗粒层)也存在很大的不一致,相应区域的功能也存在系统上的差异。研究者推断多样化的受体表达可以赋予一个区域更强的灵活性,因而受体的活动可以反映不同输入信息的整合和受到不同区域神经调节物质的影响。对于这一分子多样性和协同信息加工之间存在关联的假设,结果表明冗余-协同梯度与15种不同受体类型以及3种不同层间的受体密度的多样性增加存在显著正相关。这说明更多样化的突触受体类型意味着该区域能够响应和整合更大范围的信息,并体现在该区域的协同优势更高。
综上,通过这一跨模态和跨物种的研究,研究者对大脑动态活动进行了信息分解,阐明了人脑如何逐渐演化形成了在鲁棒性和信息整合间的平衡模式。并对于脑区间相应的冗余信息和协同信息加工程度进行了定量描述,探索了其背后的结构、遗传、神经生理、代谢与分子水平的神经机制。
