原文链接:https://mp.weixin.qq.com/s/rKH08AKypnZthAcYPiNxuw
悦影科技-赵宗亚 脑之说
背景:神经连接组理论在系统神经科学领域的广泛应用,促进了我们对人类大脑皮层拓扑组织的认识。然而,功能性连接体在小脑中的拓扑结构仍不清楚。
方法:收集1416例健康成人的静息状态功能连接(rs-fcMRI)数据。在样本1 (n = 976)中,对功能性小脑连接组的体素级和节点级拓扑特性进行了估计。此外,还分别研究了基于网络的小脑和大脑-小脑拓扑映射的拓扑性质。考虑到神经群体的时间特性,我们进一步利用隐马尔可夫模型(HMM)来揭示小脑功能连接组的动态模式。为了测试我们发现的稳健性,我们在一个独立的数据集(样本2;n = 440)运行。结果:我们发现小小脑脚I和小小脑脚II在小脑功能连接体中表现出显著的高度中心性(DC)。此外,小脑功能连接组甚至嵌套网络型小脑连接组的组织方式也表现出明显的小世界、模块化和层次化特征。小脑功能连接组中存在注意/执行网络、默认模式网络和任务正向网络三个内在模块。此外,小世界组织与层级结构的大脑-小脑相关性亦显著。通过构建大脑-小脑拓扑图,发现额顶叶和皮层下网络在小脑中的比例高于大脑皮层(3倍)。在时间特性方面,小脑功能连接组具有高度的灵活性和模块化,但在时间动态模式上表现出高度的个体特异性差异。
结论:本研究确定了功能性小脑连接组的拓扑结构和时间层次,进一步证实了小世界组织和层次结构的大脑-小脑功能耦合的显著性。
1 简介
随着神经连接组理论和神经成像技术的巨大发展,人们对大脑功能和结构特征的认识已从定位到基于网络的复杂地形。因此,系统神经科学提出了这一观点,即高阶认知活动中的高性能在很大程度上归因于基于大脑网络的拓扑结构。然而,基于网络的研究的主流长期以来一直集中在大脑皮层,而对小脑实体的见解较少。在过去的几个世纪里,小脑机能一直被这种过时的观点所支配——小脑支配运动或运动活动。然而,最近的证据表明小脑在高阶活动或功能中发挥着突出的作用,如情绪调节、执行控制、语言处理、时间感知和语言学习。此外,已有研究表明,小脑激活异常可能是神经精神疾病的诊断生物标志物,如精神分裂症、重性抑郁障碍(MDD)和注意缺陷多动障碍(ADHD)。尽管小脑发挥了重要作用,但迄今为止,基于网络的小脑地形图对小脑的作用知之甚少。
在此,本研究旨在扩展我们对小脑功能连接体的拓扑模式和时间动态状态的认识。在样本1 (n = 976)中,在图论分析的基础上,我们采用功能连接度中心性(DC)和体素镜像同位连接(VMHC)图,以体素为基础的方案描述小脑的功能连接组。进一步,为了获得基于网络的小脑轮廓,使用Seitzman-27图谱(节点=27)进行分区。为了深入研究小脑连接体是否在拓扑结构上按子网络组织,我们使用Yeo-7和Yeo-17小脑网络功能图谱构建了嵌套的网络连接体(NoN)。然后,分别从小世界性、效率性、富俱乐部组织性、模块化、协调性、同步性和层次性等方面分析了这些功能小脑连接组的拓扑结构。考虑到大脑-小脑连接的闭环回路,这些拓扑指标也被估计为大脑皮层。此外,还计算了功能性脑连接组与小脑连接组之间的跨个体拓扑耦合。同样,为了探究拓扑空间中是否存在脑-小脑耦合,我们计算了大脑和小脑连接组中每个节点的度中心性degreecentrality, DC),并以此建立拓扑连接组。最后,采用赢者通吃(WTA)策略将大脑皮层拓扑连接体网络映射到小脑。此外,为了进一步确定小脑中隐藏的时间组织类型,还对功能连接组进行了隐马尔可夫模型(HMM)。为了测试重现性,我们分析了一个独立的数据集(样本2,n = 440),以便于交叉验证我们的发现。
考虑到对p-hacking和test-retest reliability的日益关注,我们坚持APA(美国心理协会)的建议,在开放框架科学(OFS,欢迎访问:https://osf.io/zcahs/)知识库中为本研究的主要目的和目的提供预注册。
2 方法简述
为了满足日益增长的对神经成像研究透明度和可重复性的要求,本研究中使用的数据分析管道和相应脚本以标准化的方式进行了记录,方法是使用R Studio (http://projecttemplate.net/index.html)中实现的ProjectTemplate包。所有分析的日志文件在补充信息中。此外,本研究高度重视由人脑成像组织(OHBM)创建的MRI研究数据分析与共享最佳实践委员会(COBIDAS),以提高透明度和再现性。
2.1 被试
本研究使用两个独立的样本,在不同的数据点收集。样本1包括来自数据站点1的1016名参与者(678名女性,Mage: 18.70, s.d.s age: 1.28,范围:14-31),而样本2包括来自数据站点2的447名参与者(293名女性,Mage:20.13, s.d.s age: 1.68,范围:1730)。67人因数据质量问题被排除再后续分析中。
所有参与者均无神经精神病史或颅脑外伤史,并通过简易精神状态检查(MMSE)进一步证实。此外,在精神病学诊断的后验访谈中对DSM-IV的I轴和II轴进行了调查,并再次确认了他们的报告。
2.2 预处理
鉴于对头-运动擦洗策略的争论仍在继续,目前的研究努力通过结合ICA-AROMA算法和逐帧位移(FD)函数对头-运动伪影进行更严格的校正。
功能fMRI数据的预处理过程与以往关于脑功能连接组生成的研究高度平行。包含在DAPBI工具箱(http://rfmri.org/dpabi)中的fMRIPrep (V1.3.0)管道被用于功能fMRI数据的稳健自调整预处理。
值得注意的是,这里使用的是小波变换的3D去噪,而不是传统的高斯核对神经图像进行平滑处理。
2.3 功能连接组构建
为了获得均匀的功能信号,我们采用Seitzman-27图谱来预定义整个小脑节点。Seitzman图谱将整个大脑划分为300个区域,包括27个小脑区和273个大脑区 (见图1A)。此外,为了构建嵌套的网络连接组(NoN),使用了Yeo-7图谱来定义网络节点。具体来说,图谱中的每个网络(如默认模式、显著性和视觉)被定义为一个节点(见图1B)。每个参与者将生成具有7个节点的功能性连接体。为了验证这些结果是否因分区化而有偏倚,也采用了Yeo-17图谱。
为了定义功能连接体中的边,首先估计每个节点内体素的平均时间序列。然后,我们利用皮尔逊积差相关获得预定义的在小脑的每一对节点之间的相关系数来识别边,在每个小脑功能的连接体N节点有k边(k = N (N - 1)/ 2)。每个参与者的功能连接体包括N个节点和对应的k条边(见图1A)。同样,大脑连接组的边也以同样的方式估计。
图论分析为将功能连接组描述为数学模型提供了一种富有洞察力的工具,并使我们能够阐明功能连接组的拓扑性质。为此,所有连接体通过使用5%-50%的稀疏带绝对地从连接矩阵转化为二进制邻接实体。随后,分别估计了这些连接体的拓扑属性,包括效率、富俱乐部组织、模块化、关联性、同步性和层级性(见SI Method和图1C)。
鉴于大脑-小脑之间的紧密联系,本研究进一步估计了参与者的大脑-小脑拓扑耦合。具体来说,首先对每个参与者的小脑连接组和大脑连接组的拓扑属性(如小世界性)进行估计。进一步,计算所有参与者之间的Pearson相关性,以检验大脑-小脑拓扑耦合(见图1D)。
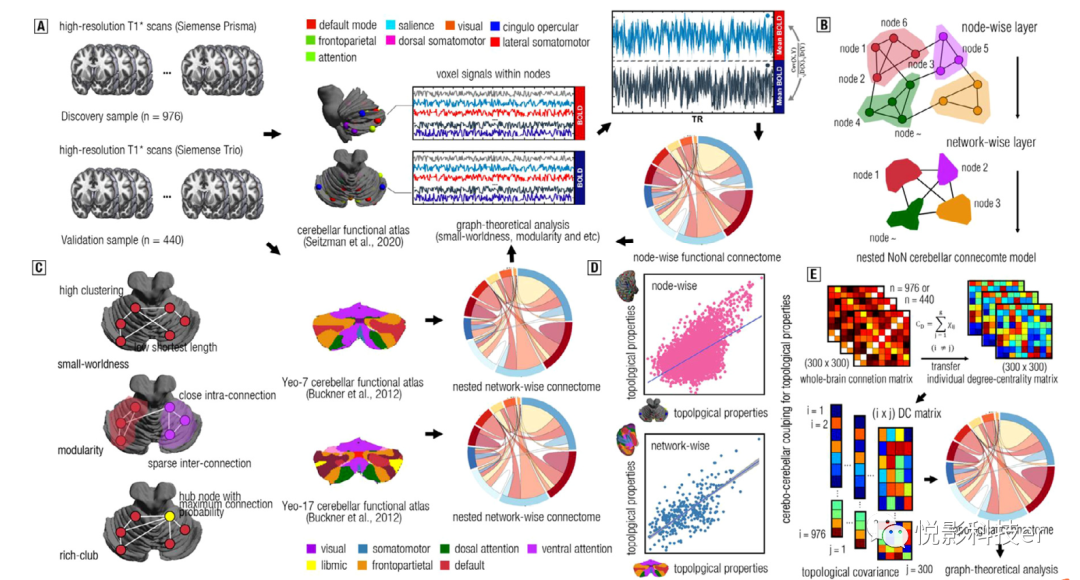
图1 当前研究的工作流程和分析管道
探讨拓扑空间中是否存在显著的大脑-小脑协方差。通过使用图论模型,计算每个参与者整个大脑中每个节点的度中心性(即表示网络中给定节点的重要性)。随后,这些度中心性之间的相关性被估计为边,每个大脑区域被用作节点来构建脑小脑连接体的拓扑连接体。此外,拓扑连接体的连接强度将用于确定哪个脑小脑连接是枢纽边。最后,我们采用赢者通吃(WTA)策略,将皮层拓扑连接体网络映射到小脑。
HMM指的是一种关于大脑自发激活的数学假设,即时间序列可以利用有限数量的隐藏状态序列进行建模。换句话说,该模型允许特定时间点(t)的多变量高斯分布(μk)的时间序列数据,从而允许观测隐藏状态(St)。对于脑功能连接组中的μk,在数学上可以将其描述为一个向量中的平均BOLD信号。除均值分布外,一旦状态k被激活,给出对应的M(通道数)xN(通道数)的方差-协方差矩阵(k):因此,HMM允许我们确定在功能性小脑连接组中是否存在固有的大脑激活的非随机的时间动态组织。
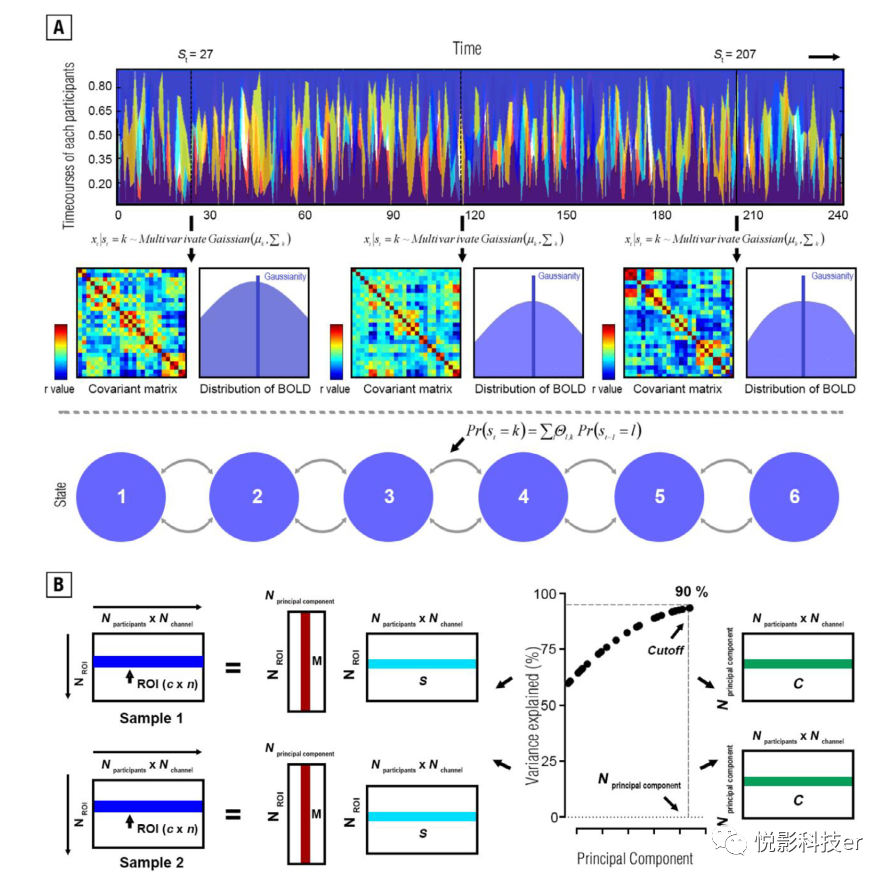
图2 隐马尔可夫模型图
3 结果
3.1 功能性小脑连接组的构建具有较高的可靠性
为了验证这些功能连接组的可靠性,本研究招募了两个大样本,即发现样本(sample 1, n = 976)和验证样本(sample 2, n = 440)。应用Seitzman图谱,通过连接图图直观检查小脑功能连接体。结果显示样本1和样本2中构建的功能性小脑连接组具有一致的模式,表明连接组构建具有较高的可靠性(见图3A)。此外,利用零秩Pearson相关分析和基于传递熵的互信息(MI, h)算法,验证了这些连接组在两个独立样本之间的再现性。结果显示,样本1和样本2的连接组强度显著相关 (图3B)。通过Jeffreys-Zellner-Siow Bayesian推断,结果显示样本1和样本2之间小脑连接组强度的相关性(见图3C)。同样,非连接组的可靠性也在Yeo-7和Yeo-17分区体中进行了检验。结果显示两种样本在非强度方面的一致性。总的来说,这些发现为验证小脑功能连接体构建的可靠性和可重复性提供了证据。
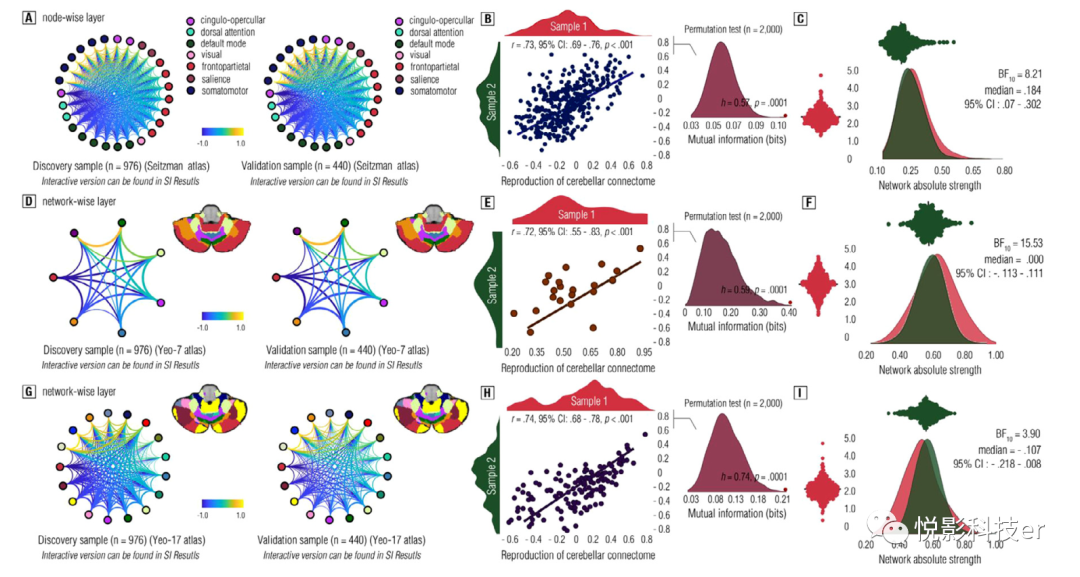
图3 小脑功能连接体重建结果
3.2 小小脑脚I和小小脑脚II是功能性小脑连接体的中枢
本研究的一个主要目标是证明功能连接体中小脑的中枢。为此,我们不仅采用绝对阈值带的二值邻接矩阵,而且采用基于体素的功能连接加权矩阵来重建小脑功能连接组,以获得无偏性的结果。结果表明,双侧小小脑脚I和小小脑脚II在功能性小脑连接组中具有显著的高度中心性(DC)(见图4A, B)。此外,这些发现在样本2中高度平行,在双侧小小脑脚I和小小脑脚II显示显著的高DC(见图4C, D),这表明高测试重测可靠性。
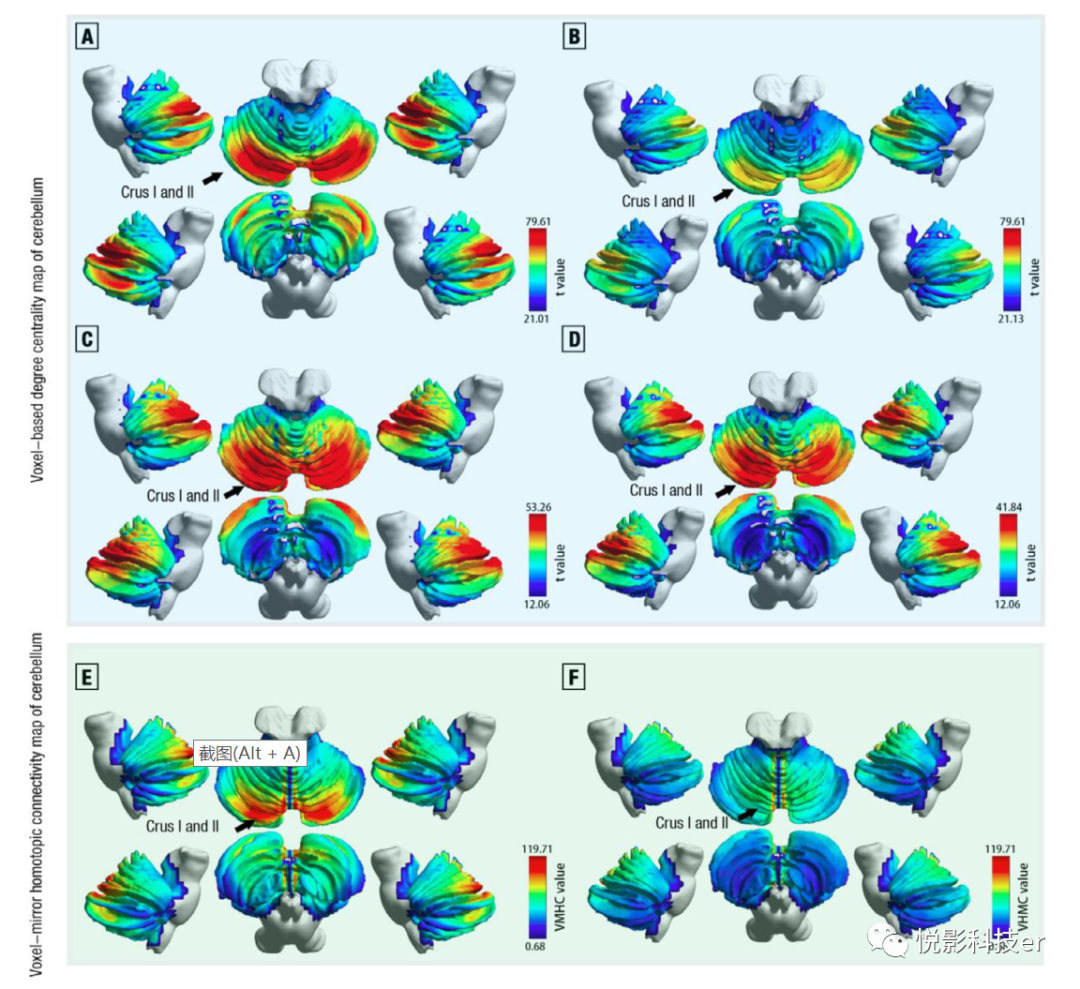
图4 在度中心性(A-D)和体素-镜像同位连通性(E-F)中基于体素的拓扑属性的脑图(t检验统计量)
体素-镜像同位连接(VMHC)描述了每个体素与其对应的大脑区域之间的功能连接,反映了大脑功能过程的同步性。本研究的另一个主要发现是在双侧小小脑脚I和小小脑脚II处观察到高VMHC (见图4E, F)。简而言之,双侧Crus I和Crus II可能是功能性小脑连接体的拓扑中心。
3.3 功能性小脑连接体以小世界、分层、模块化的方式组织
为了阐明功能性小脑连接组的拓扑性质,估计了小脑连接组的小世界组织、效率、模块性、层级性、协调性、同步性和富俱乐部结构。结果显示,无论使用何种二元阈值,发现样本和验证样本的功能性小脑连接组中都存在显著的小世界组织 (见图5A和SI表S1-2)。利用信息流理论模型,发现在两个样本中,功能性小脑连接组均具有较高的全局和局域网络效率 (见图5B, C和SI表S3 6)。总之,这些发现提供了令人信服的证据,以得出结论,功能性小脑连接组以小世界的方式组织。
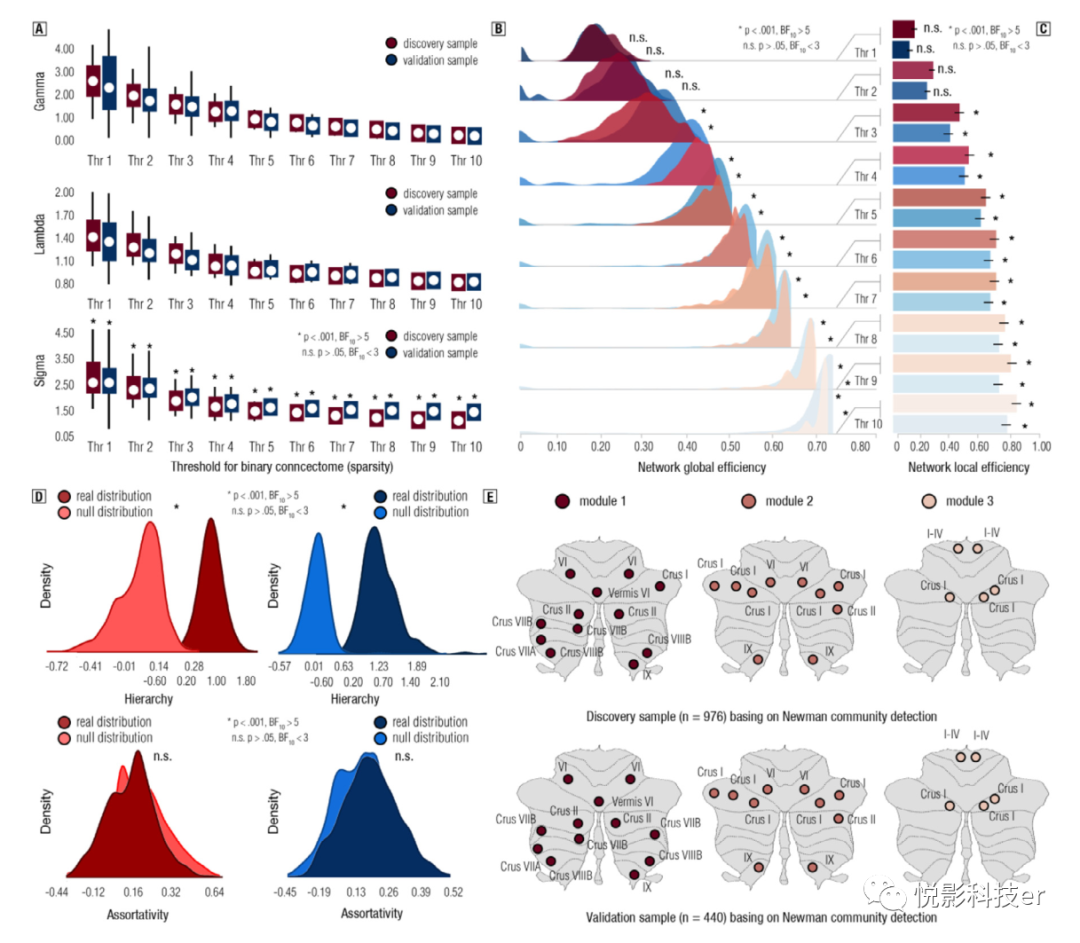
图5 小脑功能连接组的小世界组织、网络效率、层级、同配性和模块化的结果
3.4 功能性小脑连接组的NoN呈小世界和等级社区模式
我们建立了小脑的嵌套式智能网络连接组(NoN)来阐明网络到网络实体的拓扑性质。结果表明,两组样本的小脑功能网络(如视觉网络、显著网络和默认网络)均以小世界和层级方式进行组织。为了消除网络大小带来的偏差,采用Buckner(2011)功能性小脑图谱(即Yeo-17图谱)重建NONs。通过在两个样本中使用不同的网络分类方法。未发现非连接组的其他显著拓扑特性,如同步性和关联性(参见SI表S21 28)。综上所述,这些发现为小脑连接组的拓扑组织提供了有力证据,不仅是区域的拓扑组织,而且是功能模块的拓扑组织。
3.5 小世界组织和层级结构存在显著的大脑-小脑耦合
鉴于大脑-小脑连接存在闭环回路,本研究试图阐明功能连接体的大脑-小脑拓扑耦合。结果显示功能性大脑连接组与小脑连接组在小世界和层级上存在显著相关 (见图6A, B)。这些发现也在独立样本中得到验证 (见图6C, D)。因此,这些证据通过显示小世界和层级中突出的拓扑耦合,扩展了我们对大脑-小脑连接组拓扑模式的理解。也就是说,这可能表明大脑和小脑连接组在拓扑空间上存在着闭环的功能交流。
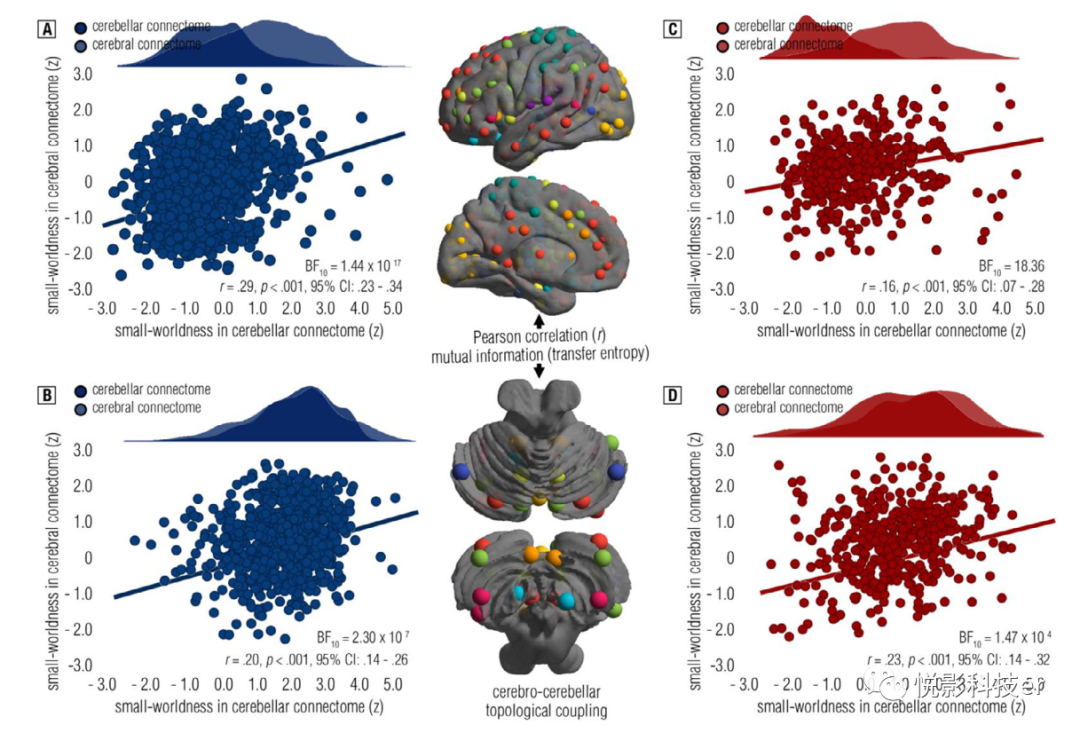
图6 小脑和大脑皮层之间拓扑耦合的散点图
3.6 拓扑协变网络提供了大脑与小脑连接体之间的拓扑映射
如前所述,对所有参与者估计了代表功能网络(连接组)重要性的每个区域的程度中心性(DC)。在此基础上,利用DC建立了跨所有参与者的拓扑协方差网络。高DC(5%)被发现在somatomotor网络(SMN),听觉网络(AN),默认模式网络(DMN),背侧注意网络(DAN)和cingulo-opercular网络(CON)(见表2和图7 a, B)。另外,结果显示了显著的强拓扑背SMN,DMN,DAN的协方差(参见图7)。社区检测分析发现六组拓扑协方差网络,包括SMN模块、DMN模块、视觉模块、任务相关模块、注意与奖励模块、皮层下与小脑模块(见图7C)。进一步,通过赢家通吃算法,将小脑连接组定义的每个节点映射到大脑连接组的固有功能网络中。结果如图7所示,显示了大部分脑-小脑映射在额顶叶网络(FPN)/皮层下网络(SCN)和小小脑脚I/II(见图7D)。小脑的FPN和SCN区分别是大脑皮层的3倍多。未展开的结果见附录一。综上所述,本研究首次构建了拓扑协方差网络,展示了脑-脑拓扑连接体和图谱。
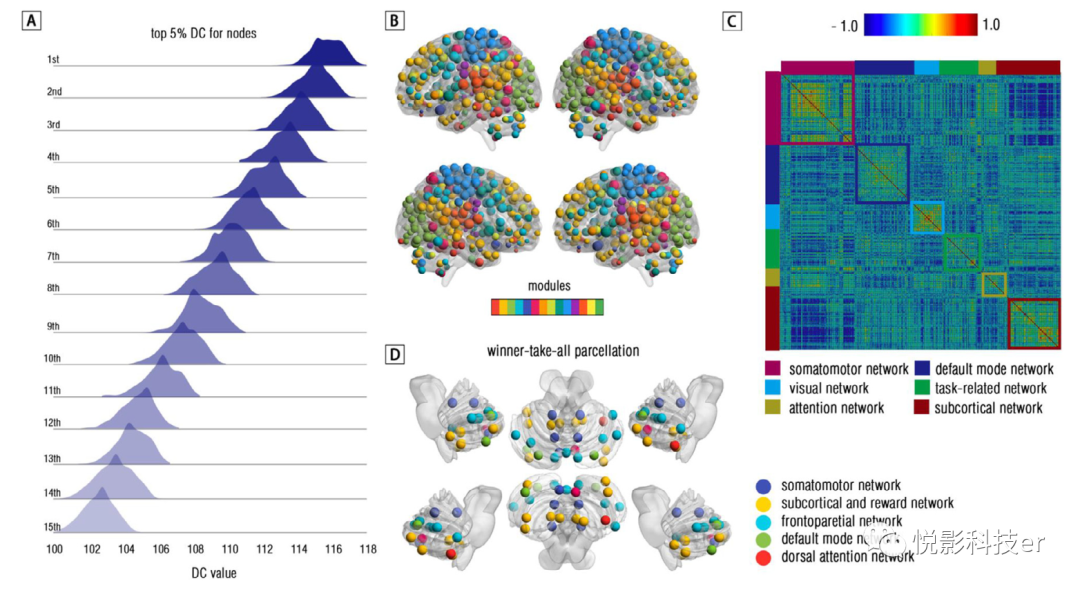
图7 由节点度中心性(DC)估计的拓扑协方差网络结果和赢家通吃算法衍生的小脑映射
3.7 小脑功能连接体的动态转换是随机和模块化的
隐马尔可夫模型(HMM)状态反映了在多个时间点重复的动态成分(集群)。在本研究中,利用变分贝叶斯推断(VBI)作为初始转弯参数,预先确定了12个隐藏状态(见材料与方法章节)。为了避免过拟合,所有预处理信号都被映射到主成分分析(PCA)空间中(64个分量,解释方差=0.902)。在多元高斯模型的基础上,发现活动对各隐态的平均模式具有显著的独立性,证明了推断状态数的合理性(见图8A)。
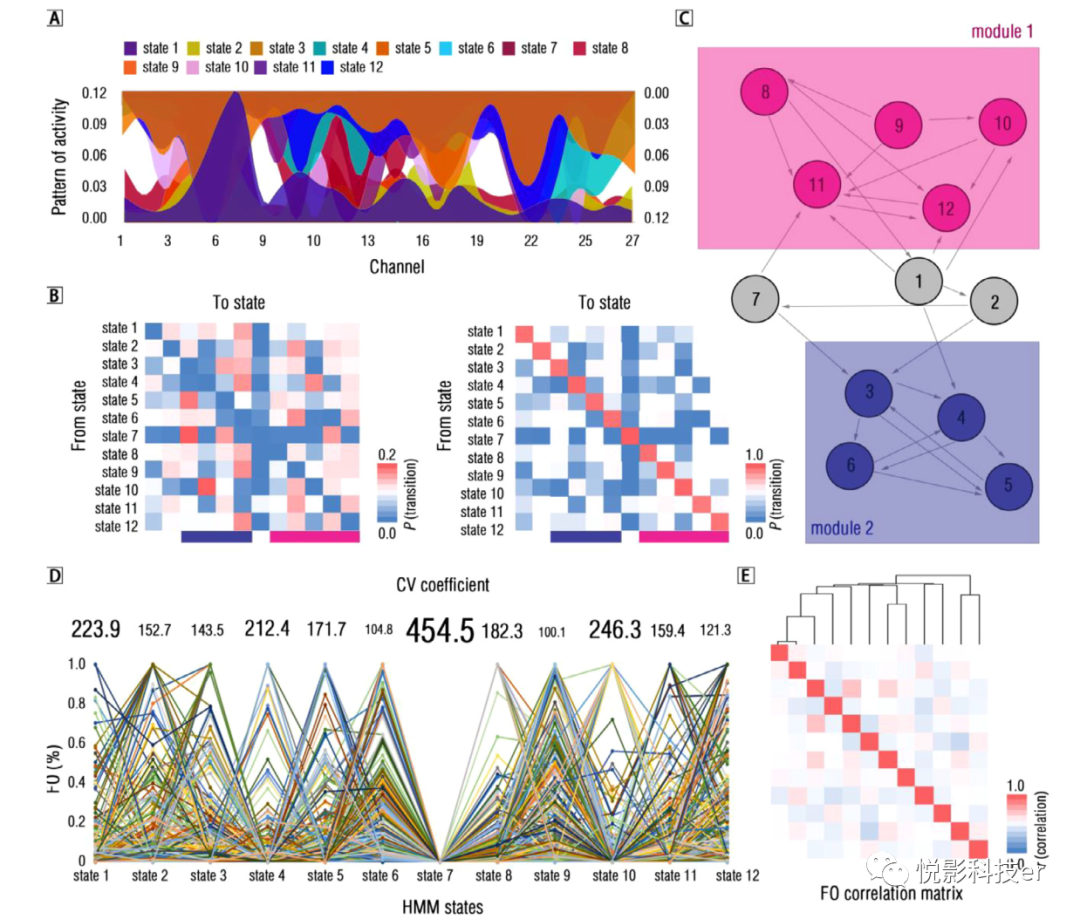
图8 功能小脑连接体的隐马尔可夫模型研究结果
3.8 每个隐藏状态所花费的时间显示了小脑连接组的高度个体特异性的时间模式和非结构动力学
除了转换概率之外,还提出了另一个量化功能性小脑连接组时间模式的指标,即占用率(FO)。它指的是花费在每个隐藏状态的时间的绝对比例。在这方面,FO提供了一种量化一个隐藏状态包含多少时间点的方法,从而便于确定隐藏状态的分配。FO的结果显示,几乎所有隐藏状态都存在显著的个体间差异,这表明了高度随机的时间模式(见图8D)。
4 讨论
本研究的主要目的是揭示小脑功能连接体的拓扑结构和时间结构。为此,我们分别采用了功能小脑连接组和嵌套网络连接组的图论模型。结果表明,两种连接体均具有显著的小世界结构、层级组织和模块化方式。此外,还发现了这些拓扑性质的大脑-小脑拓扑耦合,包括小世界和层次结构。为了进一步扩展我们对脑-小脑拓扑映射的理解,我们采用赢家通吃算法进行小脑拓扑分区。结果显示了从额顶叶网络/皮层下网络到小小脑脚I/II的显著映射。此外,HMM状态的切换是随机的,但对小脑功能连接体来说是高度模块化的。最后,小脑连接组的时间动力学已被揭示为个体内的高变异性和拓扑结构的缺乏。本研究对小脑功能连接组的拓扑结构提供了全面的认识,并为功能性小脑连接组的时间景观研究提供了新的思路。
5 结论
综上所述,本研究利用图论模型阐明了功能性小脑连接组的拓扑性质,并通过HMM模型进一步阐明了小脑的时间结构。结果表明,功能性小脑连接组的中枢为小小脑脚I和小小脑脚II。功能性小脑连接组,甚至网络型小脑连接组均表现出明显的小世界组织和层级结构。此外,在功能性小脑连接组中还观察到3个亚网络,包括注意/执行网络、默认模式网络和任务正向网络。由于脑-小脑解剖回路紧密,本研究明确了脑-小脑拓扑耦合与映射的重要意义。值得注意的是,小脑比大脑皮层表现出约3个代表额顶叶网络和皮层下网络的区域。最后,功能性小脑连接组被发现构成灵活和模块化的时间动力学。总之,本研究更详细地阐明了小脑连接组的拓扑结构和时间结构,并提出了小脑具有高性能拓扑结构,但可能受到大脑-小脑相互制约的新见解。
参考文献:Functional connectome of human cerebellum
