原文链接:https://zhuanlan.zhihu.com/p/453512950
撰文丨 @东华君
责编丨西早

长久以来,人类一直对自身所拥有的智能(或者表述为高级认知功能)引以为傲并充满兴趣。然而,受制于科学技术的发展,我们对这些能力的系统性研究开始得非常晚。大约在19世纪,人类才了解到大脑才是与我们一系列高级认知功能相关的器官,是我们“灵魂”的居所(详见前文:叩开前额叶皮层(PFC)研究的大门)。而直到上世纪70十年代末,专门研究认知历程的生物学基础的学科——认知神经科学才诞生。
直到今天,认知功能的研究已得到快速发展,然而某些错误的观念不知何时却已根深蒂固地存在于大众脑海:人类具备无语伦比的智能,并且人类的大脑皮层最为发达——因此,高级的认知功能产生于大脑皮层。那么,高级认知功能是否就是大脑皮层所产生的呢?
一、认知功能与大脑结构
随着研究的深入,我们越来越认识到:大脑是一个复杂的系统网络,许多认知功能产生和维持于这些网络中。事实上,我们在日常生活中执行的绝大多数的行为,小到伸手取物,大到吟诗作赋,都需要在大量脑区的协同作用下才能完成(图 1)。这里面既包括广受大家追捧的大脑皮层,也包括许多鲜为人知的皮层下结构。皮层下结构数量之多,分布之复杂,即便是专业的科研人员(比如我自己 )也列举不出所有的皮层下结构及其在脑中的位置。受制于技术手段和研究理念,当下绝大多数的神经科学研究往往也只局限在单个,或者少数几个脑区内。
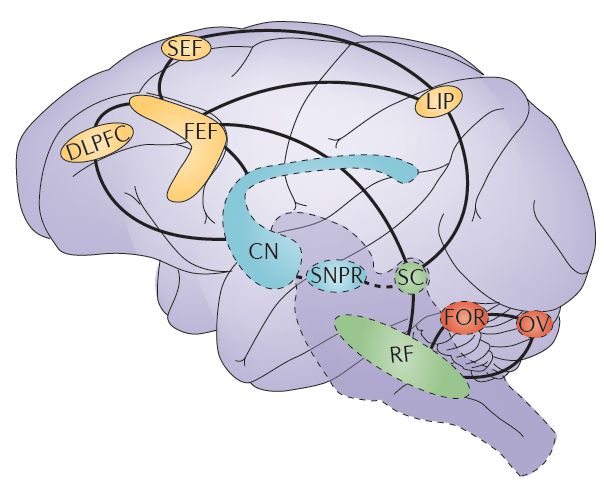
图 1. 即便是我们平常察觉不到的微眼跳(microsaccade,即眼球无意识的运动)也涉及大量的皮层和皮层下脑区之间的联动[1]。
智能不只源于大脑皮层的论述也得到了其他领域研究的验证。比如行为学的研究发现,不少智能并不是人类所特有的能力。人类平常用来标榜自己“聪明”的一些认知能力,包括记忆、计划、合作、算术运算、欺诈、创造力,甚至文化的传播都已经在大量的动物身上发现了(人类智力是对社会生活的适应,还是进化的副产品?)。这里面的某些动物甚至连大脑都没有,更遑论大脑皮层了[2]。这直接证明了很多认知功能的执行不只需要大脑皮层,甚至不需要大脑皮层也行。
二、皮层下结构参与更底层的信息处理
事实上,近些年关于认知功能的某些研究结论可能会让许多圈外人士大跌眼镜。这里面包括:一直被认为主导运动平衡的小脑其实也广泛地参与认知功能[3];我们肠道中的菌群失调也能导致认知功能障碍[4]。在最新的大量研究中,有一类我们既熟悉又陌生的研究尤其突出,正在不断地刷新我们的认知:学习的神经环路机制。
自上世纪末Wolfram Schultz等人提出奖赏预测误差假说( reward prediction error hypothesis)之后,人们对学习的神经机制理解便焕然一新。以多巴胺能系统为核心的一系列脑区成为了研究热点。虽然之后的又一系列研究表明,学习中的某一类(基于模型的学习,model-based learning)可能并不需要多巴胺的参与[5],然而其涉及的环路其实也与多巴胺能系统多有重合。而在这些环路中,最核心的节点,包括中脑、纹状体、杏仁核、海马等,大多都不位于大脑皮层。
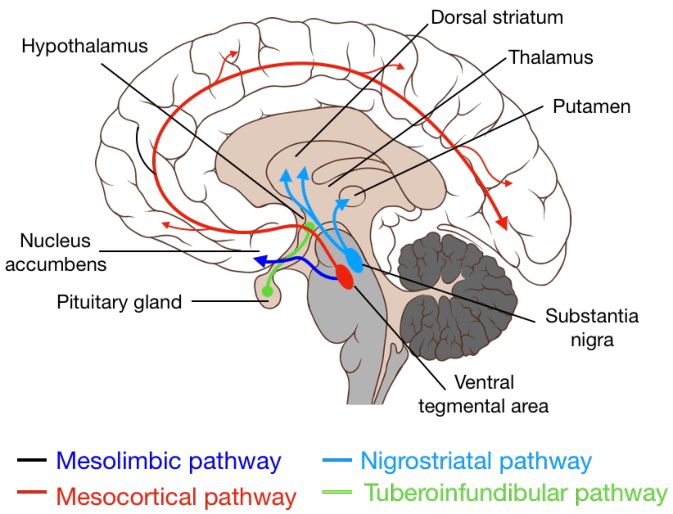
图 2.多巴胺神递质系统[6]。
三、腹侧与背侧前额叶-纹状体环路
我们实验室之前的研究主要围绕上述核团中的纹状体和杏仁核开展。简单地说,我们认为纹状体和杏仁核在强化学习中都扮演着重要的角色[7, 8]:杏仁核介导较快速的学习,纹状体介导较慢的学习;并且,腹侧纹状体和杏仁核介导强化学习中价值(value)相关信息,而背侧纹状体介导行动(action)的执行(图 3)。
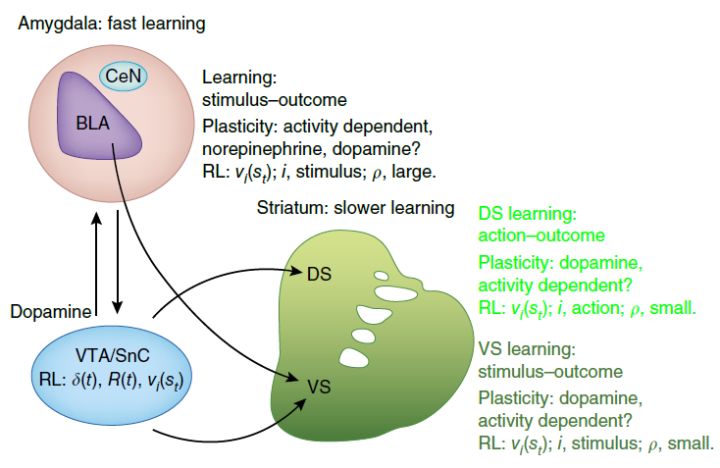
图 3. 纹状体、杏仁核和中脑之间的连接,及它们在强化学习中的角色[8]。
我的博士后课题,便是在此基础上拓展这个神经环路:研究以前额叶皮层-纹状体为核心的介导强化学习的环路机制(图 4)。我们认为,这个环路按空间位置可分为腹侧和背侧两部分[9, 10]:
腹侧环路涉及腹侧纹状体(VS)、杏仁核(BLA)、海马、腹侧苍白球(VP)、内背侧丘脑(MD)的内侧、眶额皮层(OFC)和腹内侧前额叶皮层(vmPFC),主要编码强化学习中与目标选项(包含价值、特征)相关信息。该环路服务于做出决策,即整合各种信息决定该选择哪个选项。
背侧环路涉及背侧纹状体(DS)、内侧苍白球(GPi)、内背侧丘脑(MD)的外侧、背外侧前额叶皮层(dlPFC)和后顶叶(PPC),主要介导与运动、空间位置相关的信息。该环路服务于决策的执行,即将行为决策转化为个体的动作去获取目标选项。这两条环路都受到中脑多巴胺的调节。
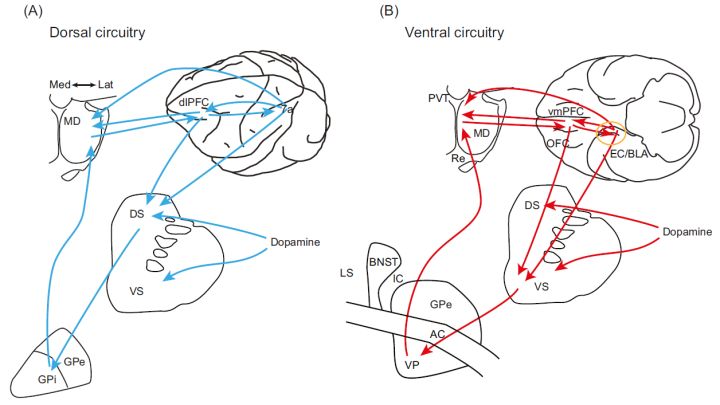
图 4.介导基于模型的强化学习的腹侧和背侧研究以前额叶皮层-纹状体神经环路[9]。
结语
除了以上举的这些例子,近些年还有大量的研究揭示了皮层下核团在我们执行认知功能时扮演了核心角色(例如,屏状核:大脑的意识开关?)。站在进化论的角度上看,或许我们的大脑皮层(特别是联合皮层)并不如我们之前认为的那么特殊。它很可能是我们所进化出的适配那些更“原始”的皮层下结构的“信号转换器”:负责将各个感觉皮层采集的信息进行整合、梳理,转换成更为简单(更低维度)的信息,转递给皮层下那些“原始”的核团使用,进而产生智能。换句话说,或许我们的大脑皮层其实只是一个兼容早期版本硬件的“华丽的适配器”(详见:人类的大脑有哪些天生的设计缺陷?;https://www.zhihu.com/question/32767604/answer/178098239)。
图片来源
其他图片:见参考文献
参考文献
1. Martinez-Conde, S., J. Otero-Millan, and S.L. Macknik, The impact of microsaccades on vision: towards a unified theory of saccadic function. Nat Rev Neurosci, 2013. 14(2): p. 83-96.
2. Bailey, C.H., et al., Is heterosynaptic modulation essential for stabilizing Hebbian plasticity and memory? Nat Rev Neurosci, 2000. 1(1): p. 11-20.
3. Carta, I., et al., Cerebellar modulation of the reward circuitry and social behavior. Science, 2019. 363(6424): eaav0581.
4. Sharon, G., et al., Human Gut Microbiota from Autism Spectrum Disorder Promote Behavioral Symptoms in Mice. Cell, 2019. 177(6): p. 1600-1618.e17.
5. Clark, J.J., N.G. Hollon, and P.E. Phillips, Pavlovian valuation systems in learning and decision making. Curr Opin Neurobiol, 2012. 22(6): p. 1054-1061.
6. Nummenmaa, L., K. Seppälä, and V. Putkinen, Molecular imaging of the human emotion circuit. 2020.
7. Costa, V.D., A.R. Mitz, and B.B. Averbeck, Subcortical Substrates of Explore-Exploit Decisions in Primates. Neuron, 2019. 103(3): p. 533-545 e5.
8. Averbeck, B.B. and V.D. Costa, Motivational neural circuits underlying reinforcement learning. Nat Neurosci, 2017. 20(4): p. 505-512.
9. Averbeck, B.B. and E.A. Murray, Hypothalamic Interactions with Large-Scale Neural Circuits Underlying Reinforcement Learning and Motivated Behavior. Trends Neurosci, 2020. 43(9): p. 681-694.
10. Neftci, E.O. and B.B. Averbeck, Reinforcement learning in artificial and biological systems. Nature Machine Intelligence, 2019. 1(3): p. 133-143.
